

Evolution of sex allocation plasticity in a hermaphroditic flatworm genus
- PMID: 35583959
- PMCID: PMC9321609
- DOI: 10.1111/jeb.14020
Evolution of sex allocation plasticity in a hermaphroditic flatworm genus
Abstract
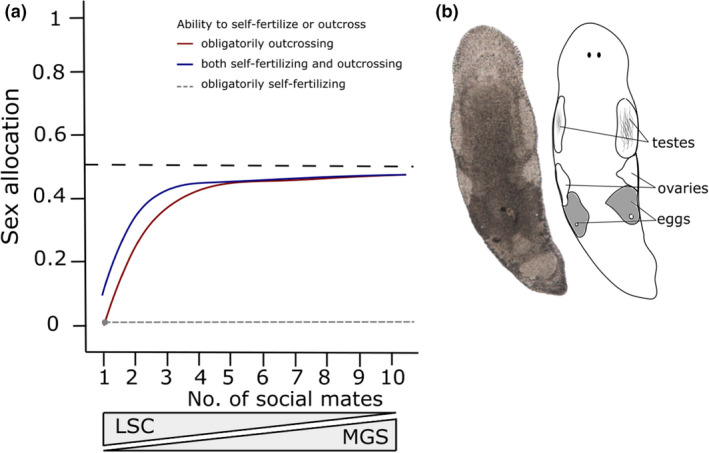
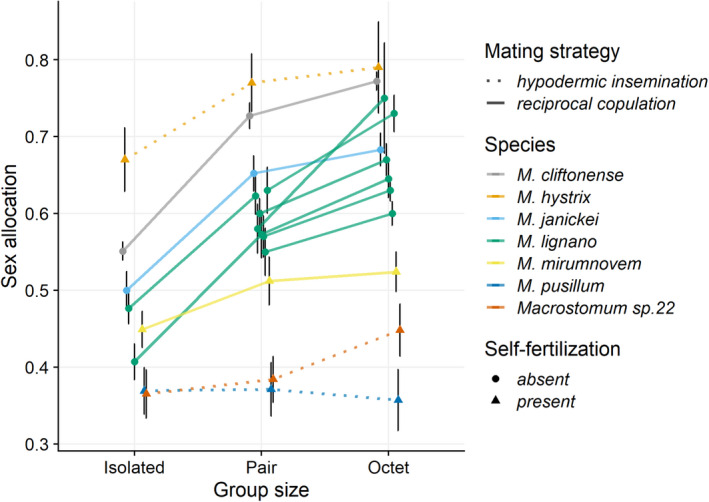
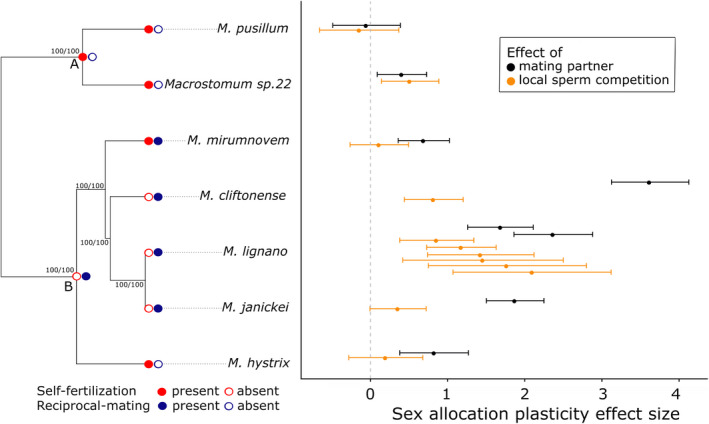
Sex allocation theory in simultaneous hermaphrodites predicts that optimal sex allocation is influenced by local sperm competition, which occurs when related sperm compete to fertilize a given set of eggs. Different factors, including the mating strategy and the ability to self-fertilize, are predicted to affect local sperm competition and hence the optimal SA. Moreover, since the local sperm competition experienced by an individual can vary temporally and spatially, this can favour the evolution of sex allocation plasticity. Here, using seven species of the free-living flatworm genus Macrostomum, we document interspecific variation in sex allocation, but neither their mating strategy nor their ability to self-fertilize significantly predicted sex allocation among these species. Since we also found interspecific variation in sex allocation plasticity, we further estimated standardized effect sizes for plasticity in response to (i) the presence of mating partners (i.e. in isolation vs. with partners) and (ii) the strength of local sperm competition (i.e. in small vs. large groups). We found that self-fertilization predicted sex allocation plasticity with respect to the presence of mating partners, with plasticity being lower for self-fertilizing species. Finally, we showed that interspecific variation in sex allocation is higher than intraspecific variation due to sex allocation plasticity. Our study suggests that both sex allocation and sex allocation plasticity are evolutionarily labile, with self-fertilization predicting the latter in Macrostomum.
Keywords: hypodermic insemination; local mate competition; phenotypic plasticity; reciprocal mating; self-fertilization; sperm competition.
© 2022 The Authors. Journal of Evolutionary Biology published by John Wiley & Sons Ltd on behalf of European Society for Evolutionary Biology.
Conflict of interest statement
The authors hereby declare no conflict of interest.
Figures
Similar articles
-
Mating behavior and reproductive morphology predict macroevolution of sex allocation in hermaphroditic flatworms.BMC Biol. 2022 Feb 7;20(1):35. doi: 10.1186/s12915-022-01234-1. BMC Biol. 2022. PMID: 35130880 Free PMC article.
-
Variation in sex allocation plasticity in three closely related flatworm species.Ecol Evol. 2019 Aug 16;10(1):26-37. doi: 10.1002/ece3.5566. eCollection 2020 Jan. Ecol Evol. 2019. PMID: 31988714 Free PMC article.
-
Self-fertilization, sex allocation and spermatogenesis kinetics in the hypodermically inseminating flatworm Macrostomum pusillum.J Exp Biol. 2017 May 1;220(Pt 9):1568-1577. doi: 10.1242/jeb.149682. Epub 2017 Feb 9. J Exp Biol. 2017. PMID: 28183866
-
Sex allocation and investment into pre- and post-copulatory traits in simultaneous hermaphrodites: the role of polyandry and local sperm competition.Philos Trans R Soc Lond B Biol Sci. 2013 Jan 21;368(1613):20120052. doi: 10.1098/rstb.2012.0052. Print 2013 Mar 5. Philos Trans R Soc Lond B Biol Sci. 2013. PMID: 23339243 Free PMC article. Review.
-
Sperm competition and the evolution of gametic compatibility in externally fertilizing taxa.Mol Hum Reprod. 2014 Dec;20(12):1190-7. doi: 10.1093/molehr/gau069. Epub 2014 Oct 16. Mol Hum Reprod. 2014. PMID: 25323969 Review.
Cited by
-
Hermaphroditic origins of anisogamy.Philos Trans R Soc Lond B Biol Sci. 2023 May 8;378(1876):20220283. doi: 10.1098/rstb.2022.0283. Epub 2023 Mar 20. Philos Trans R Soc Lond B Biol Sci. 2023. PMID: 36934747 Free PMC article.
-
Evolution and co-evolution of the suck behaviour, a postcopulatory female resistance trait that manipulates received ejaculate.BMC Biol. 2025 Mar 26;23(1):87. doi: 10.1186/s12915-025-02171-5. BMC Biol. 2025. PMID: 40140885 Free PMC article.
References
-
- Ax, P. (1951). Die Turbellarien des Eulitorals der Kieler Bucht. Zoologische Jahrbuecher Abteilung Fuer Systematik Oekologie Und Geographie Der Tiere, 80, 277–378.
-
- Ax, P. (1956). Les Turbellariés des étangs côtiers du littoral méditerranéen de la France méridionale… Hermann et Cie.
-
- Ax, P. (2008). Plathelminthes aus Brackgewässern der Nordhalbkugel. Abhandlungen der Mathematisch‐naturwissenschaftligen Klasse (pp. 696). Verlag der Akademie der Wissenschaften und der Literatur.
-
- Bates, D. , Mächler, M. , Bolker, B. M. , & Walker, S. C. (2015). Fitting linear mixed‐effects models using lme4. Journal of Statistical Software, 67, 1–48. 10.18637/jss.v067.i01 - DOI
Publication types
MeSH terms
Associated data
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
