

Furin and TMPRSS2 Resistant Spike Induces Robust Humoral and Cellular Immunity Against SARS-CoV-2 Lethal Infection
- PMID: 35585971
- PMCID: PMC9108258
- DOI: 10.3389/fimmu.2022.872047
Furin and TMPRSS2 Resistant Spike Induces Robust Humoral and Cellular Immunity Against SARS-CoV-2 Lethal Infection
Abstract
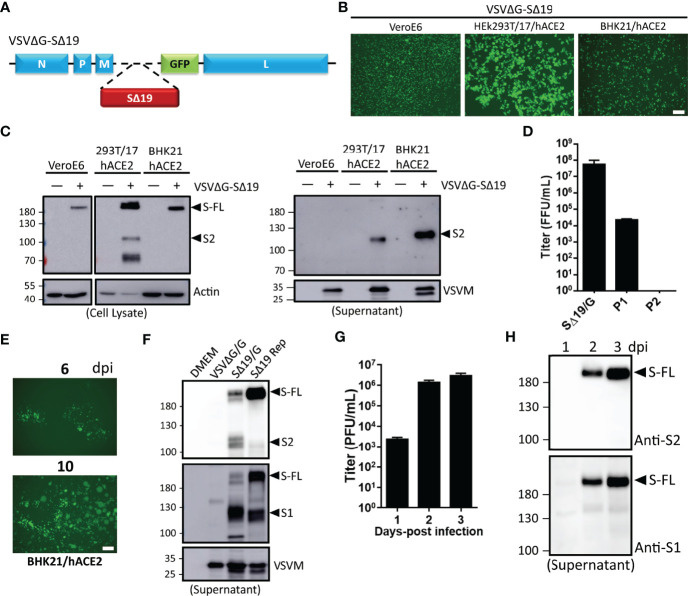
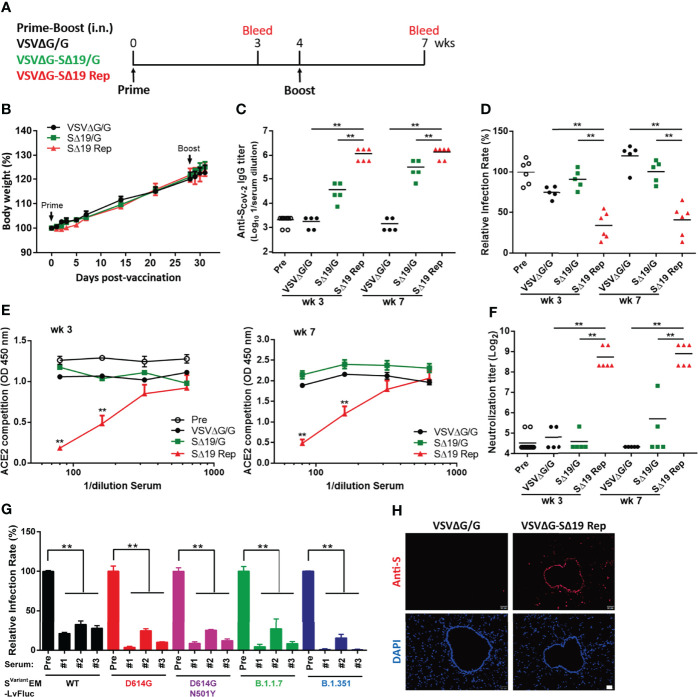
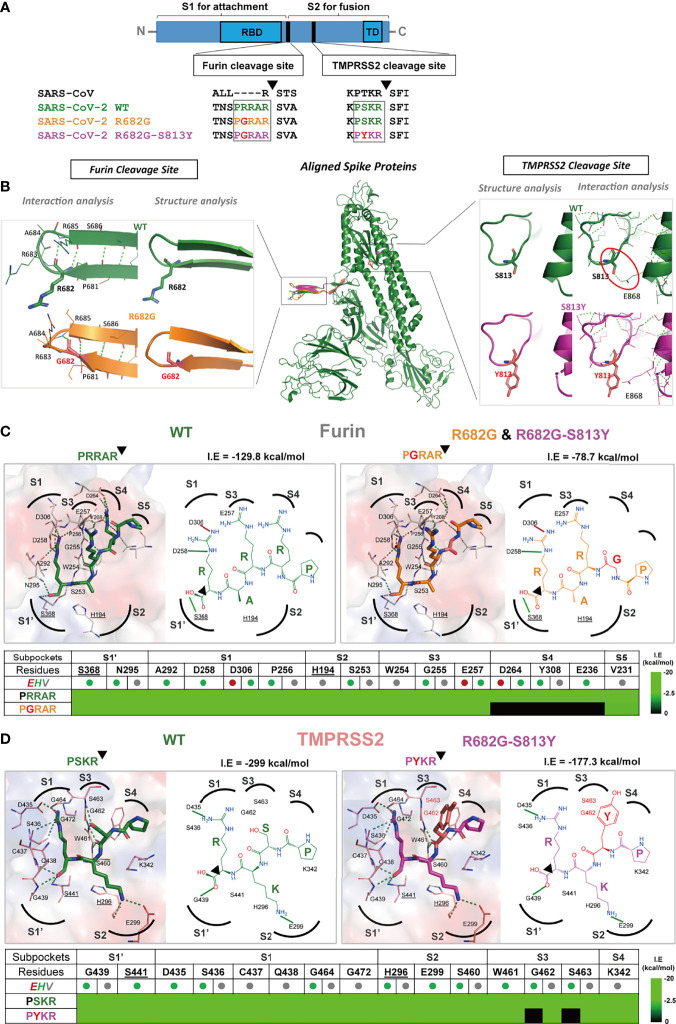
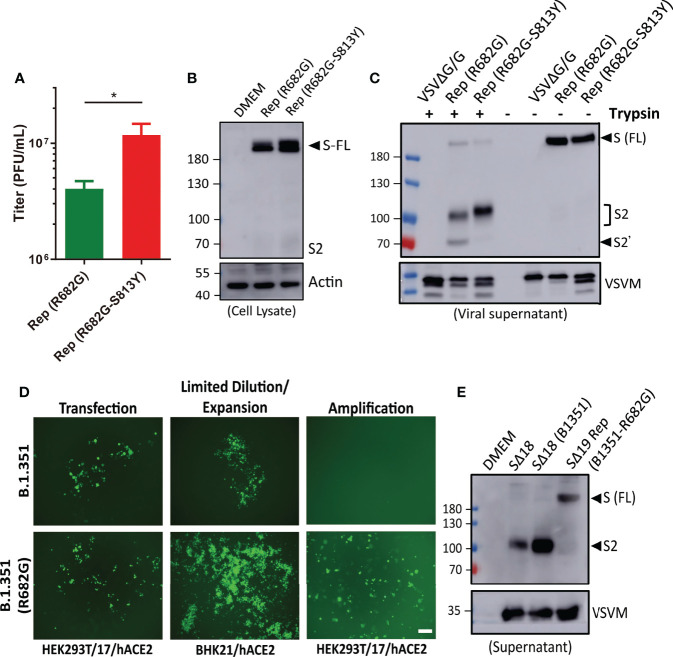
An effective COVID-19 vaccine against broad SARS-CoV-2 variants is still an unmet need. In the study, the vesicular stomatitis virus (VSV)-based vector was used to express the SARS-CoV-2 Spike protein to identify better vaccine designs. The replication-competent of the recombinant VSV-spike virus with C-terminal 19 amino acid truncation (SΔ19 Rep) was generated. A single dose of SΔ19 Rep intranasal vaccination is sufficient to induce protective immunity against SARS-CoV-2 infection in hamsters. All the clones isolated from the SΔ19 Rep virus contained R682G mutation located at the Furin cleavage site. An additional S813Y mutation close to the TMPRSS2 cleavage site was identified in some clones. The enzymatic processing of S protein was blocked by these mutations. The vaccination of the R682G-S813Y virus produced a high antibody response against S protein and a robust S protein-specific CD8+ T cell response. The vaccinated animals were protected from the lethal SARS-CoV-2 (delta variant) challenge. The S antigen with resistance to enzymatic processes by Furin and TMPRSS2 will provide better immunogenicity for vaccine design.
Keywords: ACE2 transgenic mice; S1/S2 cleavage site; SARS-CoV-2 Spike; TMPRSS2; VSV; furin; pseudotype; replication-competent.
Copyright © 2022 Lin, Tien, Kuo, Lin, Tsai, Chen, Tsai, Su, Pathak, Yang, Yu, Chuang, Wu, Tsai, Dai, Liao, Chai, Su, Chuang, Liu, Chen, Dou, Chen, Chen, Liao and Yu.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
Similar articles
-
Distinctive Roles of Furin and TMPRSS2 in SARS-CoV-2 Infectivity.J Virol. 2022 Apr 27;96(8):e0012822. doi: 10.1128/jvi.00128-22. Epub 2022 Mar 28. J Virol. 2022. PMID: 35343766 Free PMC article.
-
Reactive Centre Loop Mutagenesis of SerpinB3 to Target TMPRSS2 and Furin: Inhibition of SARS-CoV-2 Cell Entry and Replication.Int J Mol Sci. 2022 Oct 19;23(20):12522. doi: 10.3390/ijms232012522. Int J Mol Sci. 2022. PMID: 36293378 Free PMC article.
-
Rapid SARS-CoV-2 Adaptation to Available Cellular Proteases.J Virol. 2022 Mar 9;96(5):e0218621. doi: 10.1128/jvi.02186-21. Epub 2022 Jan 12. J Virol. 2022. PMID: 35019723 Free PMC article.
-
Roles of the polybasic furin cleavage site of spike protein in SARS-CoV-2 replication, pathogenesis, and host immune responses and vaccination.J Med Virol. 2022 May;94(5):1815-1820. doi: 10.1002/jmv.27539. Epub 2021 Dec 31. J Med Virol. 2022. PMID: 34936124 Review.
-
Proteolytic activation of SARS-CoV-2 spike protein.Microbiol Immunol. 2022 Jan;66(1):15-23. doi: 10.1111/1348-0421.12945. Epub 2021 Oct 12. Microbiol Immunol. 2022. PMID: 34561887 Free PMC article. Review.
Cited by
-
Spike substitution T813S increases Sarbecovirus fusogenicity by enhancing the usage of TMPRSS2.PLoS Pathog. 2023 May 17;19(5):e1011123. doi: 10.1371/journal.ppat.1011123. eCollection 2023 May. PLoS Pathog. 2023. PMID: 37196033 Free PMC article.
-
Glycosylation and S-palmitoylation regulate SARS-CoV-2 spike protein intracellular trafficking.iScience. 2022 Aug 19;25(8):104709. doi: 10.1016/j.isci.2022.104709. Epub 2022 Jul 3. iScience. 2022. PMID: 35813875 Free PMC article.
-
TLR9 and STING agonists cooperatively boost the immune response to SARS-CoV-2 RBD vaccine through an increased germinal center B cell response and reshaped T helper responses.Int J Biol Sci. 2023 May 29;19(9):2897-2913. doi: 10.7150/ijbs.81210. eCollection 2023. Int J Biol Sci. 2023. PMID: 37324951 Free PMC article.
References
Publication types
MeSH terms
Substances
Supplementary concepts
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous