

MDM2 E3 ligase activity is essential for p53 regulation and cell cycle integrity
- PMID: 35588102
- PMCID: PMC9119546
- DOI: 10.1371/journal.pgen.1010171
MDM2 E3 ligase activity is essential for p53 regulation and cell cycle integrity
Erratum in
-
Correction: MDM2 E3 ligase activity is essential for p53 regulation and cell cycle integrity.PLoS Genet. 2022 Jun 27;18(6):e1010293. doi: 10.1371/journal.pgen.1010293. eCollection 2022 Jun. PLoS Genet. 2022. PMID: 35759469 Free PMC article.
Abstract
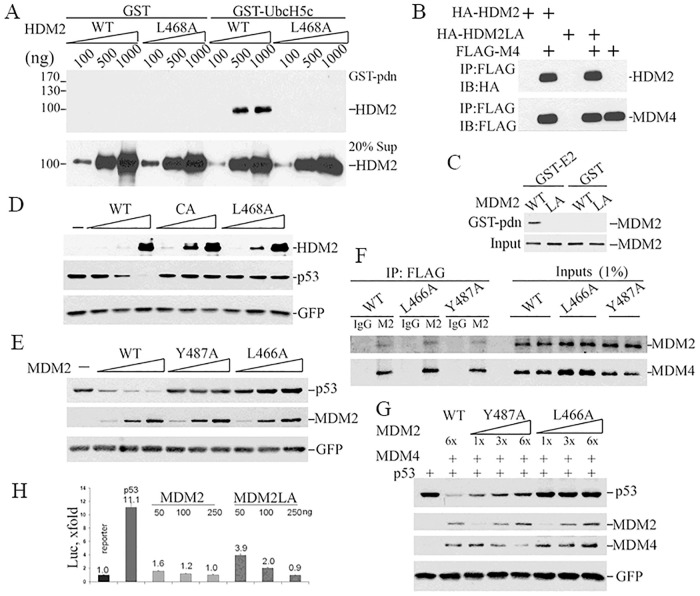
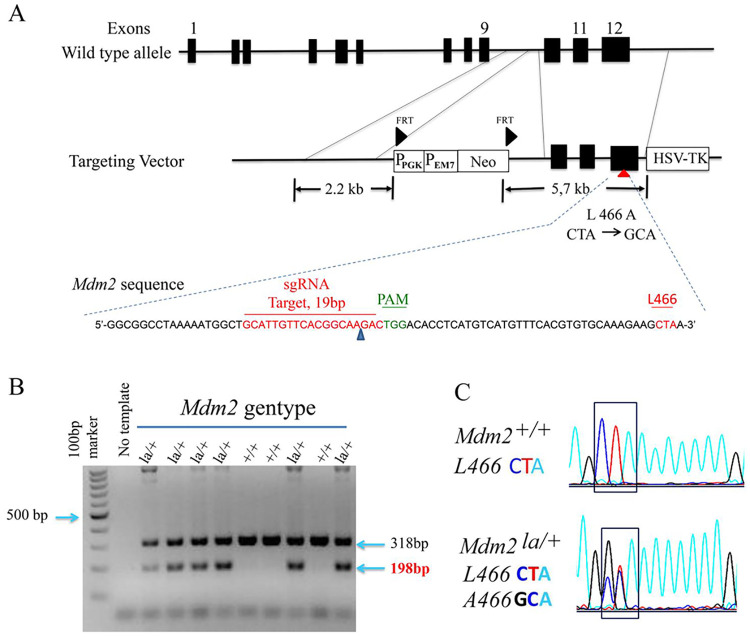
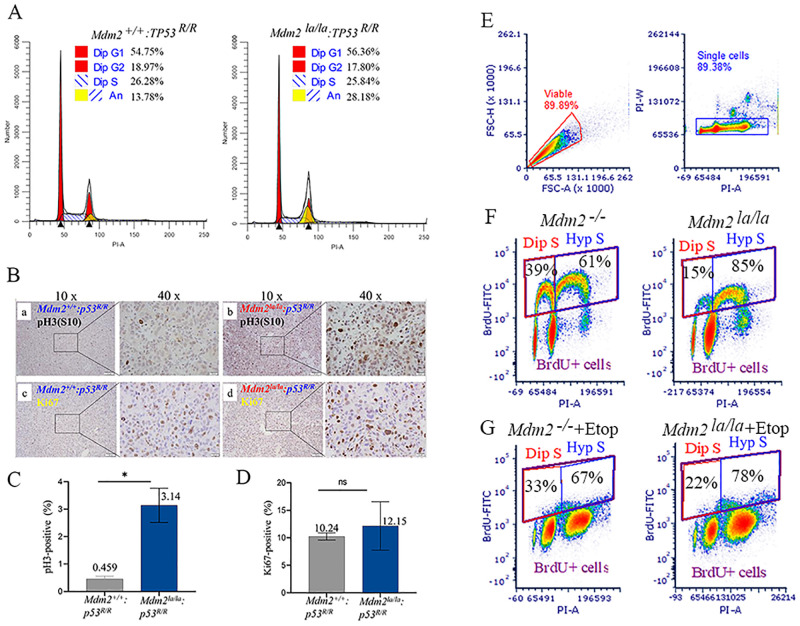
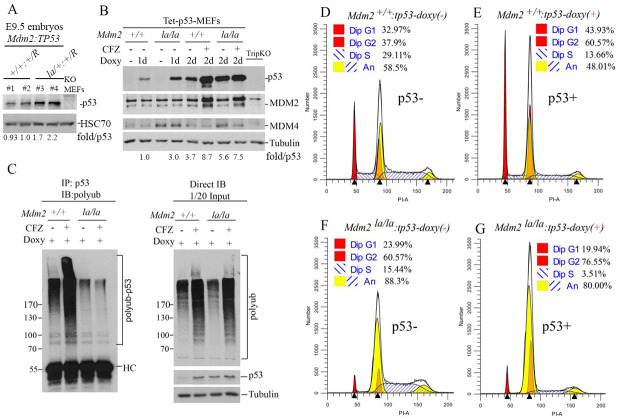
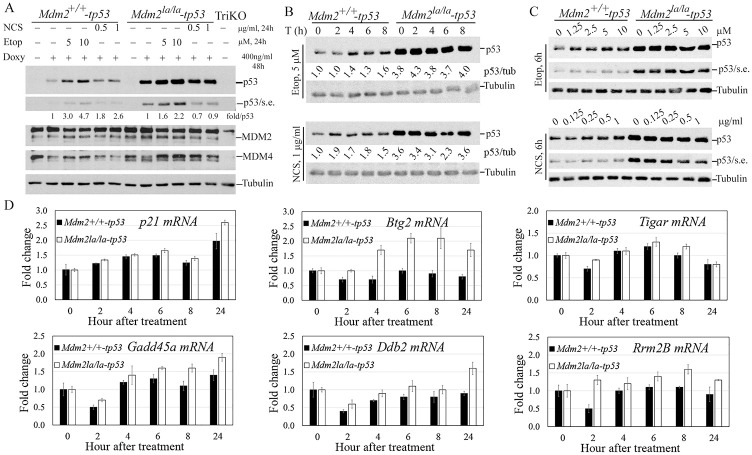
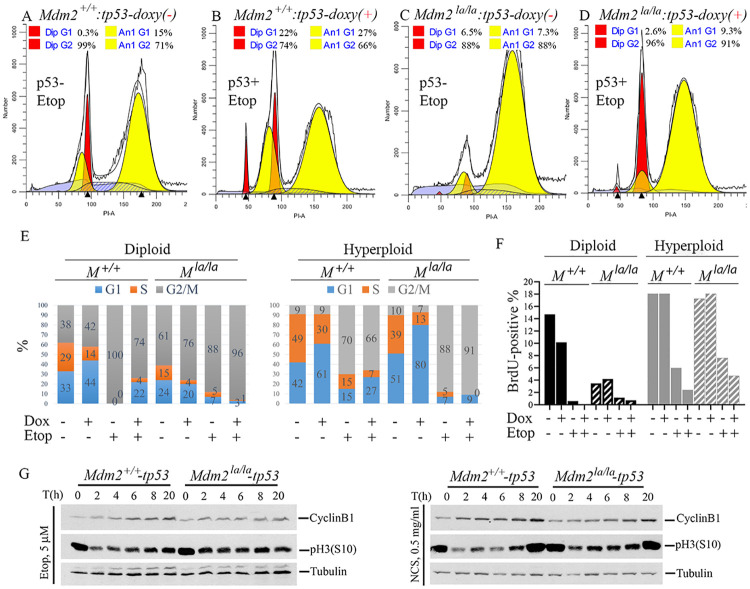
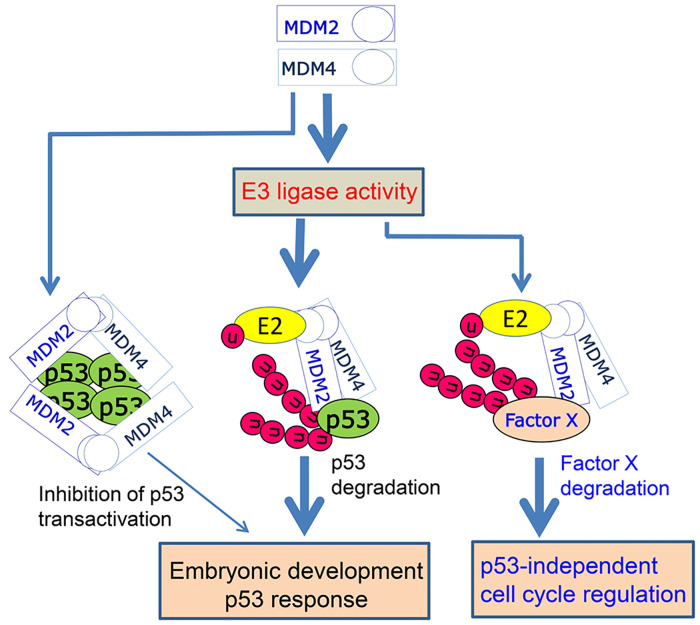
MDM2 and MDM4 are key regulators of p53 and function as oncogenes when aberrantly expressed. MDM2 and MDM4 partner to suppress p53 transcriptional transactivation and polyubiquitinate p53 for degradation. The importance of MDM2 E3-ligase-mediated p53 regulation remains controversial. To resolve this, we generated mice with an Mdm2 L466A mutation that specifically compromises E2 interaction, abolishing MDM2 E3 ligase activity while preserving its ability to bind MDM4 and suppress p53 transactivation. Mdm2L466A/L466A mice exhibit p53-dependent embryonic lethality, demonstrating MDM2 E3 ligase activity is essential for p53 regulation in vivo. Unexpectedly, cells expressing Mdm2L466A manifest cell cycle G2-M transition defects and increased aneuploidy even in the absence of p53, suggesting MDM2 E3 ligase plays a p53-independent role in cell cycle regulation and genome integrity. Furthermore, cells bearing the E3-dead MDM2 mutant show aberrant cell cycle regulation in response to DNA damage. This study uncovers an uncharacterized role for MDM2's E3 ligase activity in cell cycle beyond its essential role in regulating p53's stability in vivo.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
