

Interplay between SERCA, 4E-BP, and eIF4E in the Drosophila heart
- PMID: 35588119
- PMCID: PMC9119464
- DOI: 10.1371/journal.pone.0267156
Interplay between SERCA, 4E-BP, and eIF4E in the Drosophila heart
Abstract
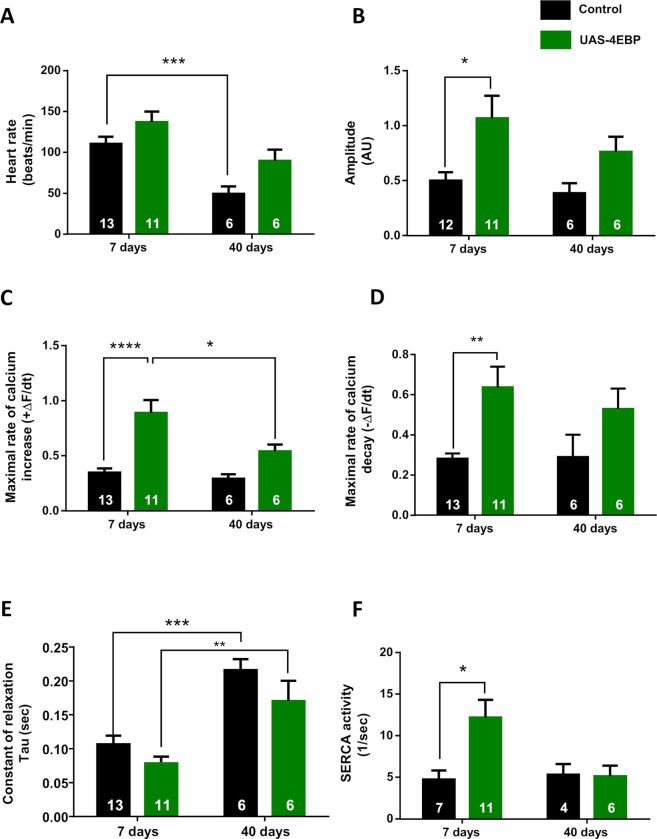
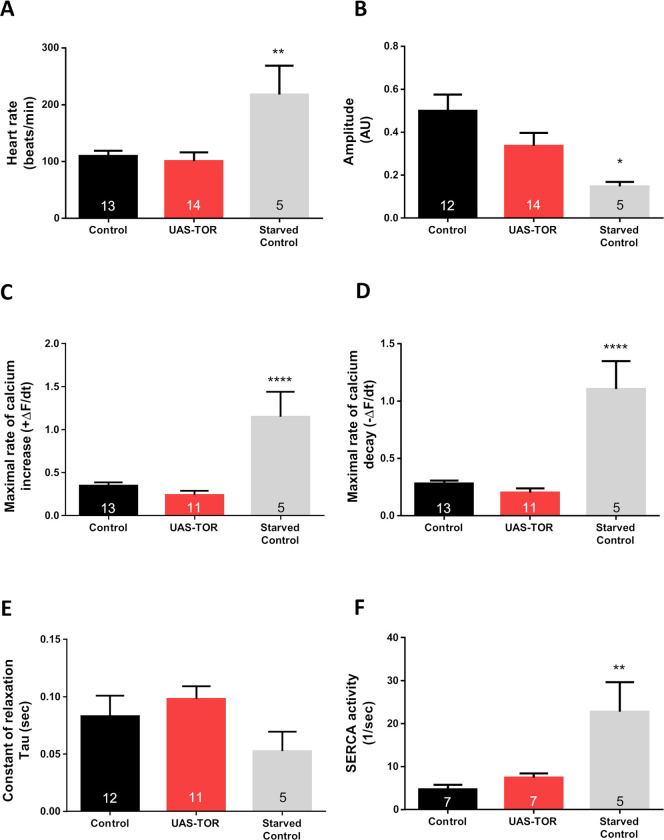
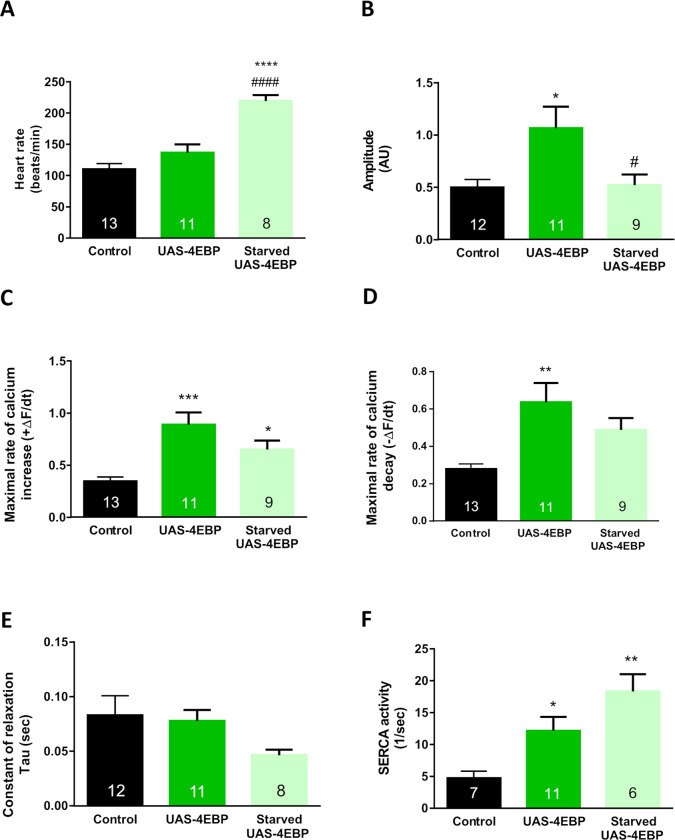
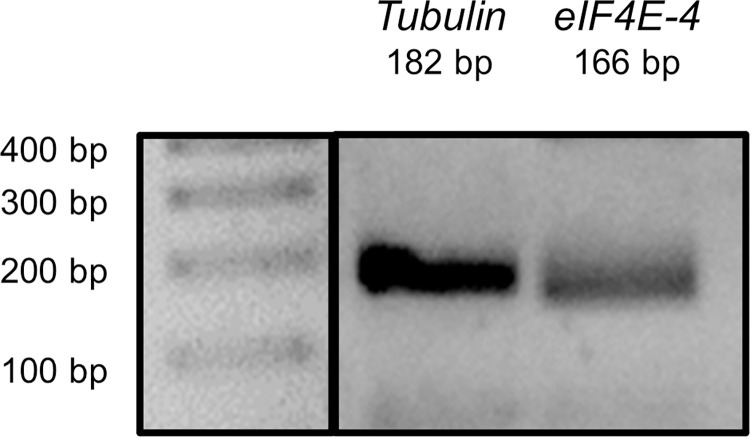
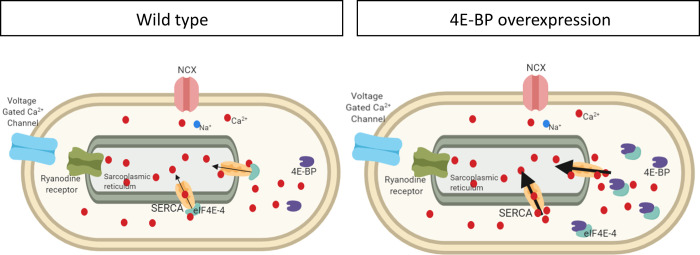
Appropriate cardiac performance depends on a tightly controlled handling of Ca2+ in a broad range of species, from invertebrates to mammals. The role of the Ca2+ ATPase, SERCA, in Ca2+ handling is pivotal, and its activity is regulated, inter alia, by interacting with distinct proteins. Herein, we give evidence that 4E binding protein (4E-BP) is a novel regulator of SERCA activity in Drosophila melanogaster during cardiac function. Flies over-expressing 4E-BP showed improved cardiac performance in young individuals associated with incremented SERCA activity. Moreover, we demonstrate that SERCA interacts with translation initiation factors eIF4E-1, eIF4E-2 and eIF4E-4 in a yeast two-hybrid assay. The specific identification of eIF4E-4 in cardiac tissue leads us to propose that the interaction of elF4E-4 with SERCA may be the basis of the cardiac effects observed in 4E-BP over-expressing flies associated with incremented SERCA activity.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures

References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
