

Templated folding of the RTX domain of the bacterial toxin adenylate cyclase revealed by single molecule force spectroscopy
- PMID: 35589788
- PMCID: PMC9120197
- DOI: 10.1038/s41467-022-30448-8
Templated folding of the RTX domain of the bacterial toxin adenylate cyclase revealed by single molecule force spectroscopy
Abstract
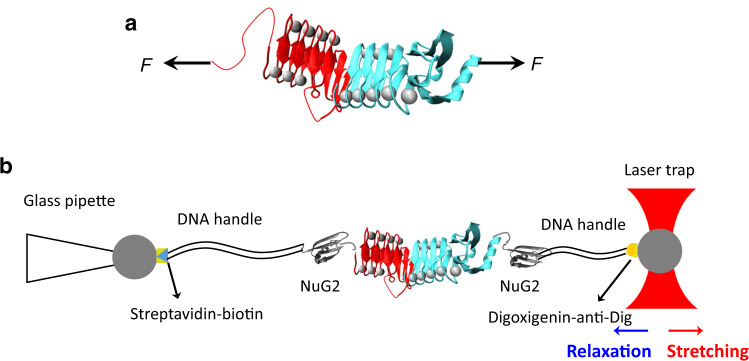
The RTX (repeats-in-toxin) domain of the bacterial toxin adenylate cyclase (CyaA) contains five RTX blocks (RTX-i to RTX-v) and its folding is essential for CyaA's functions. It was shown that the C-terminal capping structure of RTX-v is critical for the whole RTX to fold. However, it is unknown how the folding signal transmits within the RTX domain. Here we use optical tweezers to investigate the interplay between the folding of RTX-iv and RTX-v. Our results show that RTX-iv alone is disordered, but folds into a Ca2+-loaded-β-roll structure in the presence of a folded RTX-v. Folding trajectories of RTX-iv-v reveal that the folding of RTX-iv is strictly conditional upon the folding of RTX-v, suggesting that the folding of RTX-iv is templated by RTX-v. This templating effect allows RTX-iv to fold rapidly, and provides significant mutual stabilization. Our study reveals a possible mechanism for transmitting the folding signal within the RTX domain.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures





Similar articles
-
The adenylate cyclase toxin RTX domain follows a series templated folding mechanism with implications for toxin activity.J Biol Chem. 2023 Sep;299(9):105150. doi: 10.1016/j.jbc.2023.105150. Epub 2023 Aug 9. J Biol Chem. 2023. PMID: 37567473 Free PMC article.
-
Continuous Assembly of β-Roll Structures Is Implicated in the Type I-Dependent Secretion of Large Repeat-in-Toxins (RTX) Proteins.J Mol Biol. 2020 Sep 18;432(20):5696-5710. doi: 10.1016/j.jmb.2020.08.020. Epub 2020 Aug 27. J Mol Biol. 2020. PMID: 32860773
-
Characterization of the regions involved in the calcium-induced folding of the intrinsically disordered RTX motifs from the bordetella pertussis adenylate cyclase toxin.J Mol Biol. 2010 Mar 26;397(2):534-49. doi: 10.1016/j.jmb.2010.01.031. Epub 2010 Jan 22. J Mol Biol. 2010. PMID: 20096704
-
Block V RTX Domain of Adenylate Cyclase from Bordetella pertussis: A Conformationally Dynamic Scaffold for Protein Engineering Applications.Toxins (Basel). 2017 Sep 17;9(9):289. doi: 10.3390/toxins9090289. Toxins (Basel). 2017. PMID: 28926974 Free PMC article. Review.
-
Calcium-dependent disorder-to-order transitions are central to the secretion and folding of the CyaA toxin of Bordetella pertussis, the causative agent of whooping cough.Toxicon. 2018 Jul;149:37-44. doi: 10.1016/j.toxicon.2018.01.007. Epub 2018 Jan 12. Toxicon. 2018. PMID: 29337218 Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
