

Proton-gated anion transport governs macropinosome shrinkage
- PMID: 35590106
- PMCID: PMC9203271
- DOI: 10.1038/s41556-022-00912-0
Proton-gated anion transport governs macropinosome shrinkage
Abstract
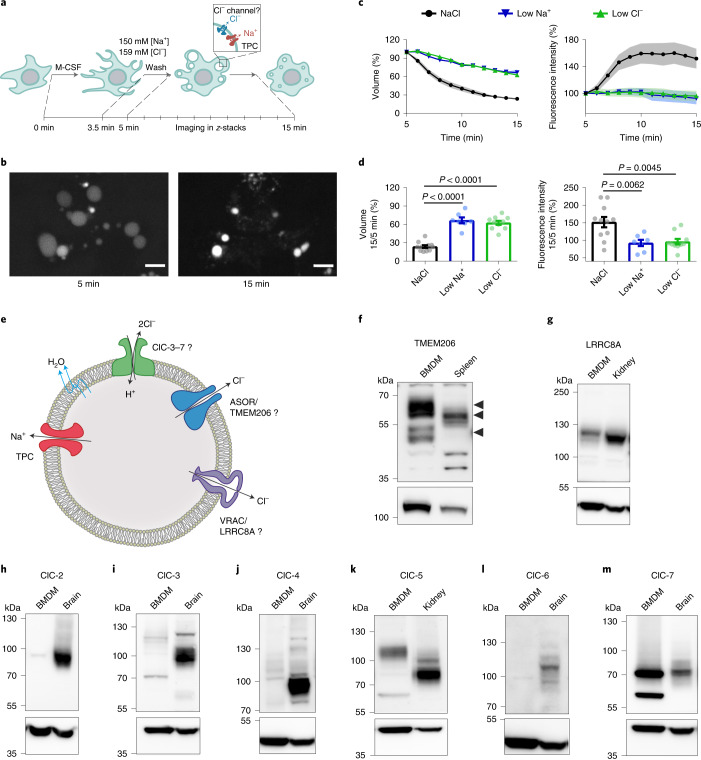
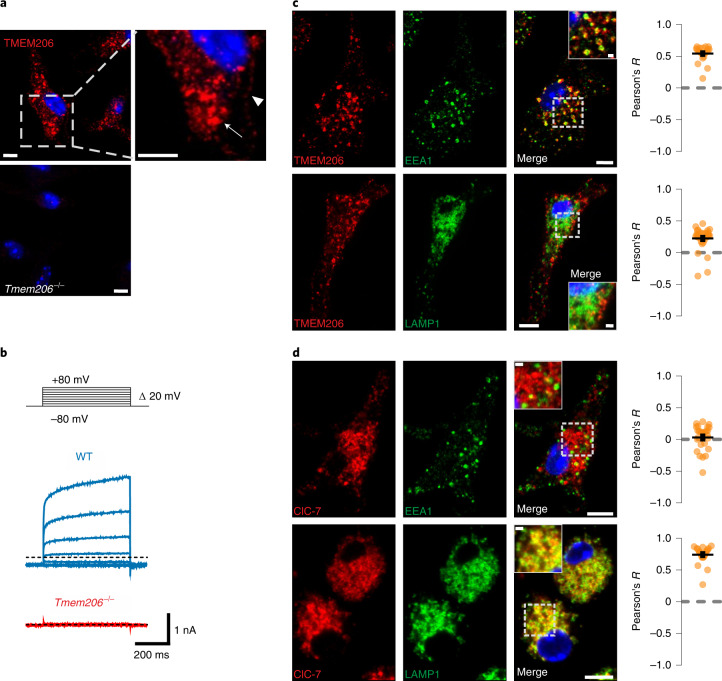
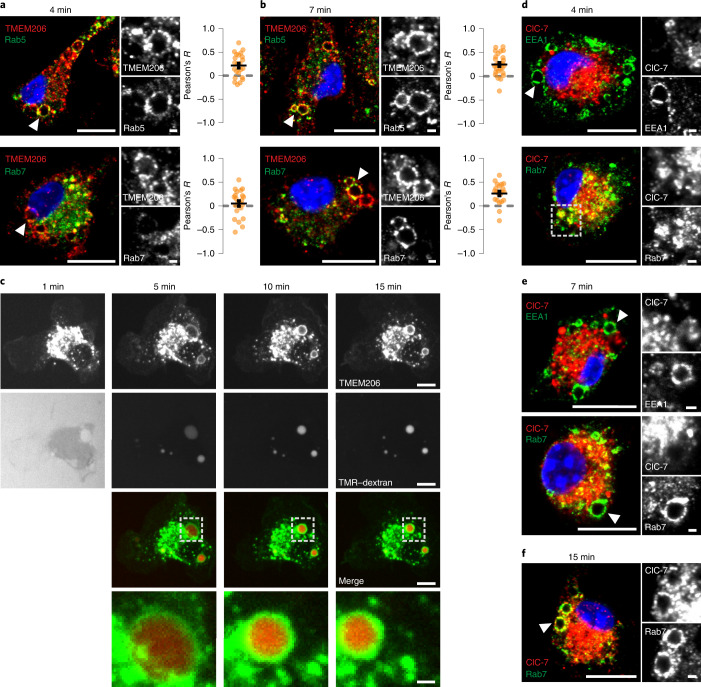
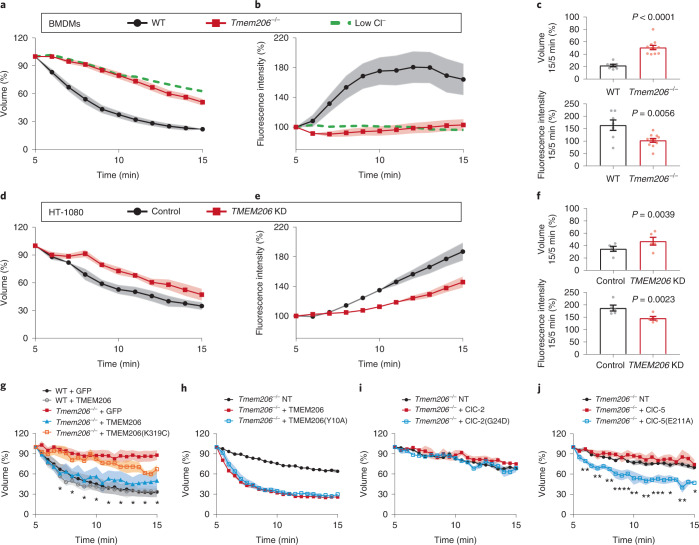
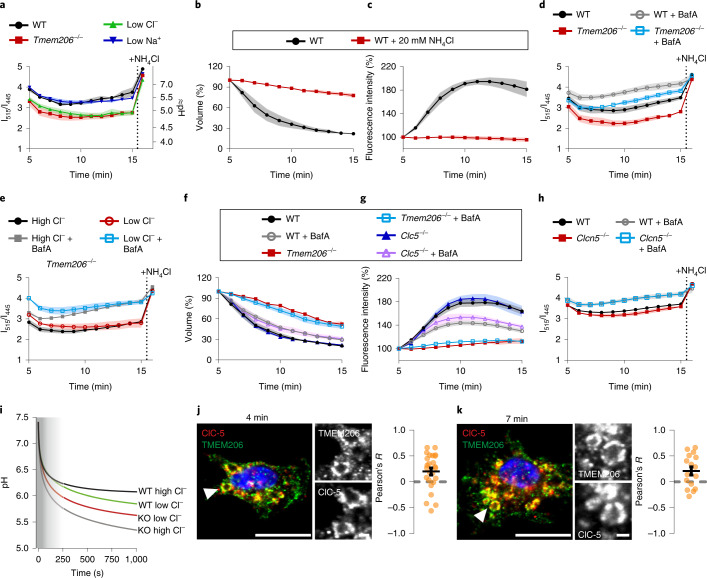
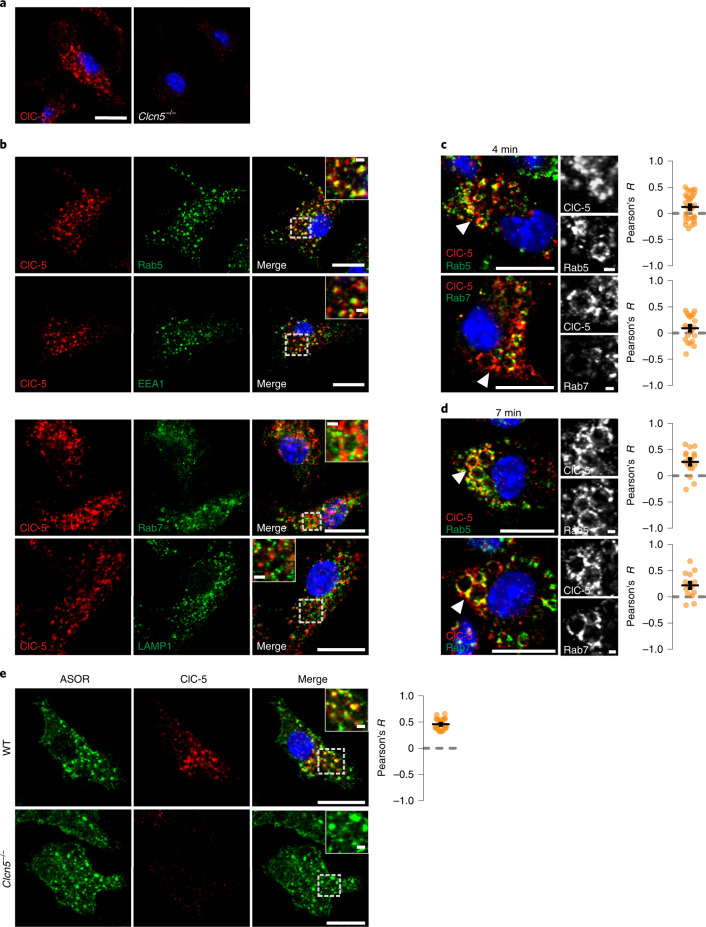
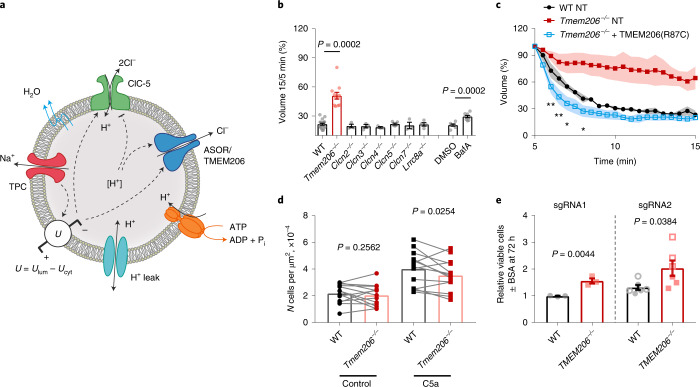
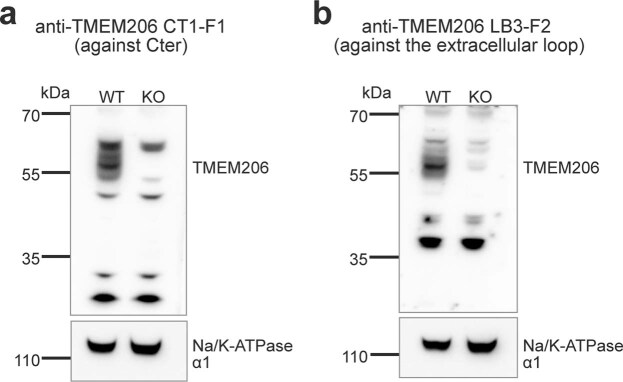
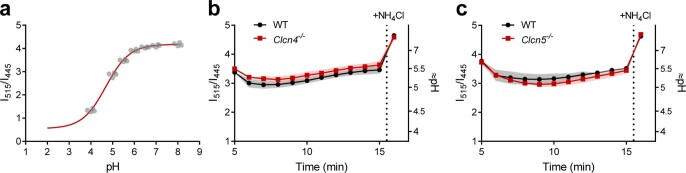
Intracellular organelles change their size during trafficking and maturation. This requires the transport of ions and water across their membranes. Macropinocytosis, a ubiquitous form of endocytosis of particular importance for immune and cancer cells, generates large vacuoles that can be followed optically. Shrinkage of macrophage macropinosomes depends on TPC-mediated Na+ efflux and Cl- exit through unknown channels. Relieving osmotic pressure facilitates vesicle budding, positioning osmotic shrinkage upstream of vesicular sorting and trafficking. Here we identify the missing macrophage Cl- channel as the proton-activated Cl- channel ASOR/TMEM206. ASOR activation requires Na+-mediated depolarization and luminal acidification by redundant transporters including H+-ATPases and CLC 2Cl-/H+ exchangers. As corroborated by mathematical modelling, feedback loops requiring the steep voltage and pH dependencies of ASOR and CLCs render vacuole resolution resilient towards transporter copy numbers. TMEM206 disruption increased albumin-dependent survival of cancer cells. Our work suggests a function for the voltage and pH dependence of ASOR and CLCs, provides a comprehensive model for ion-transport-dependent vacuole maturation and reveals biological roles of ASOR.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
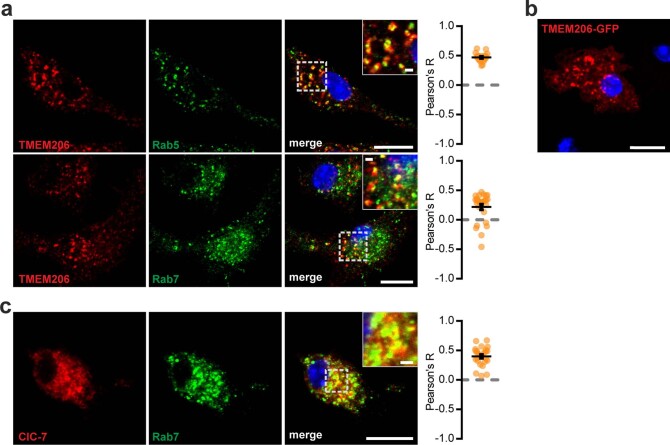
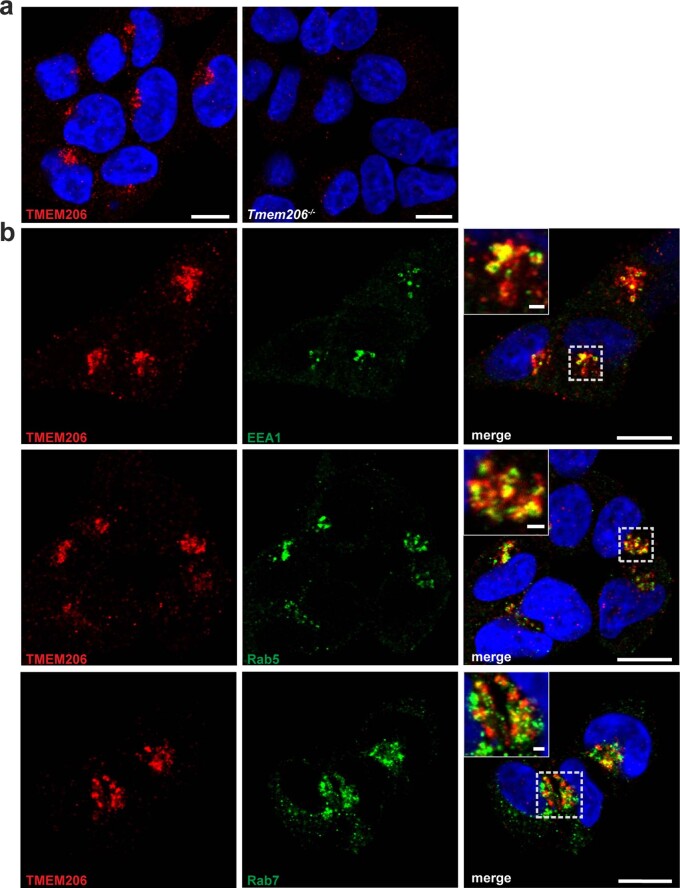
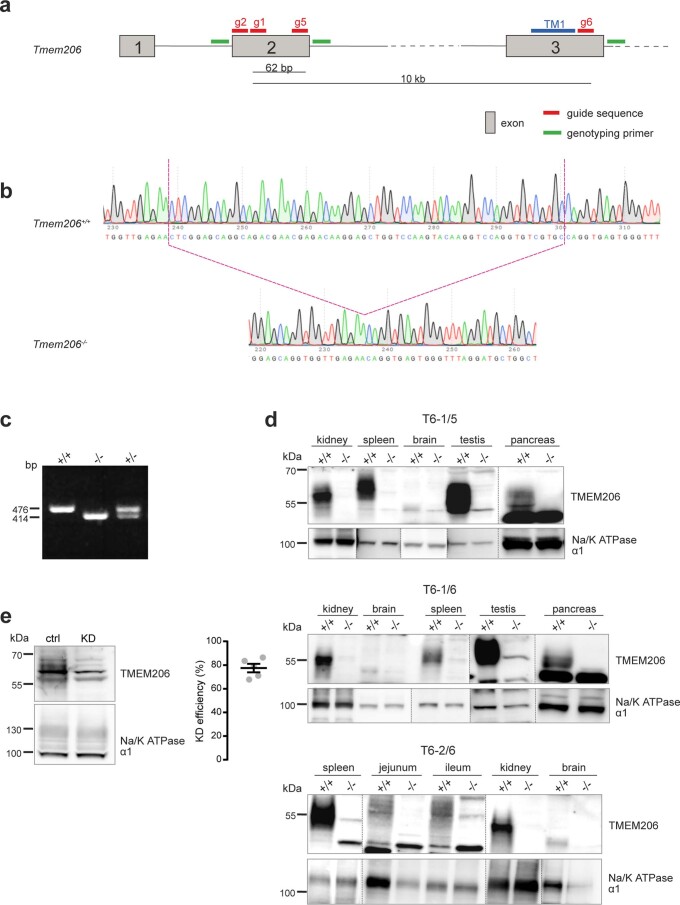
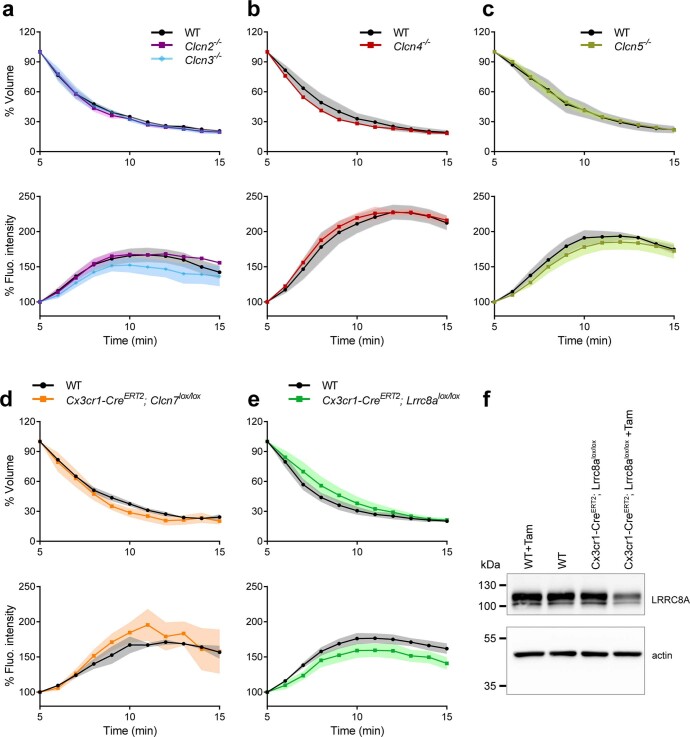
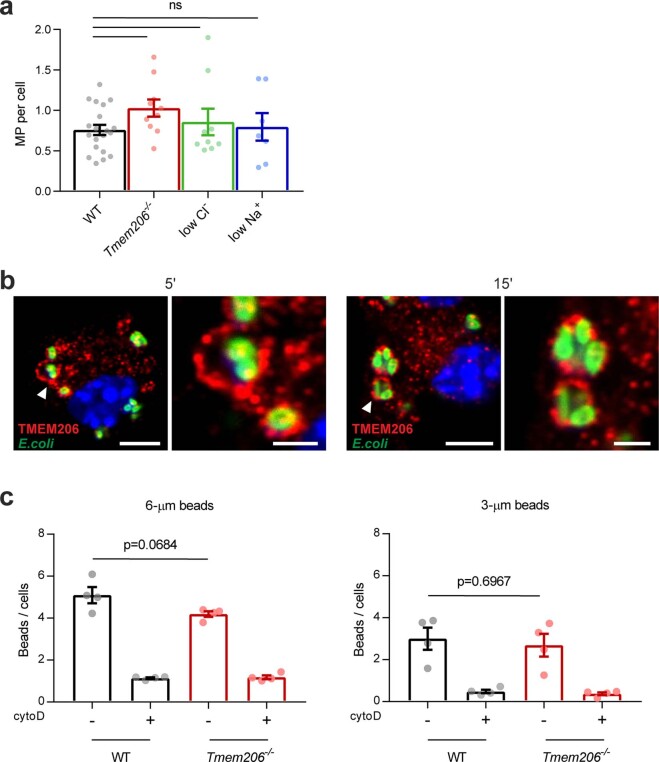
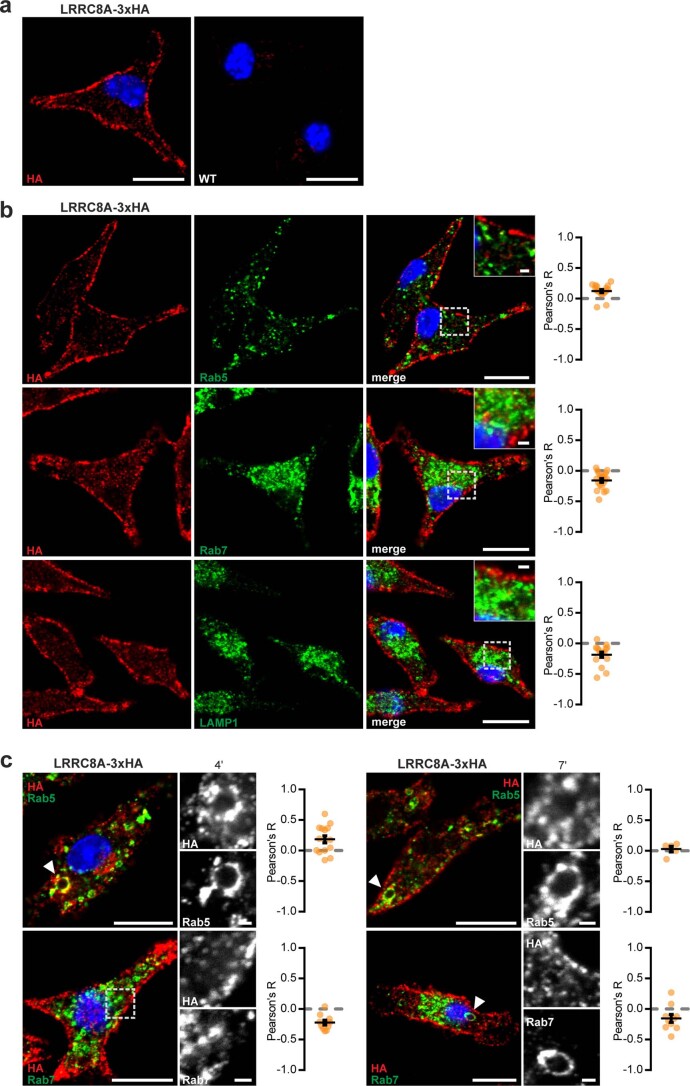
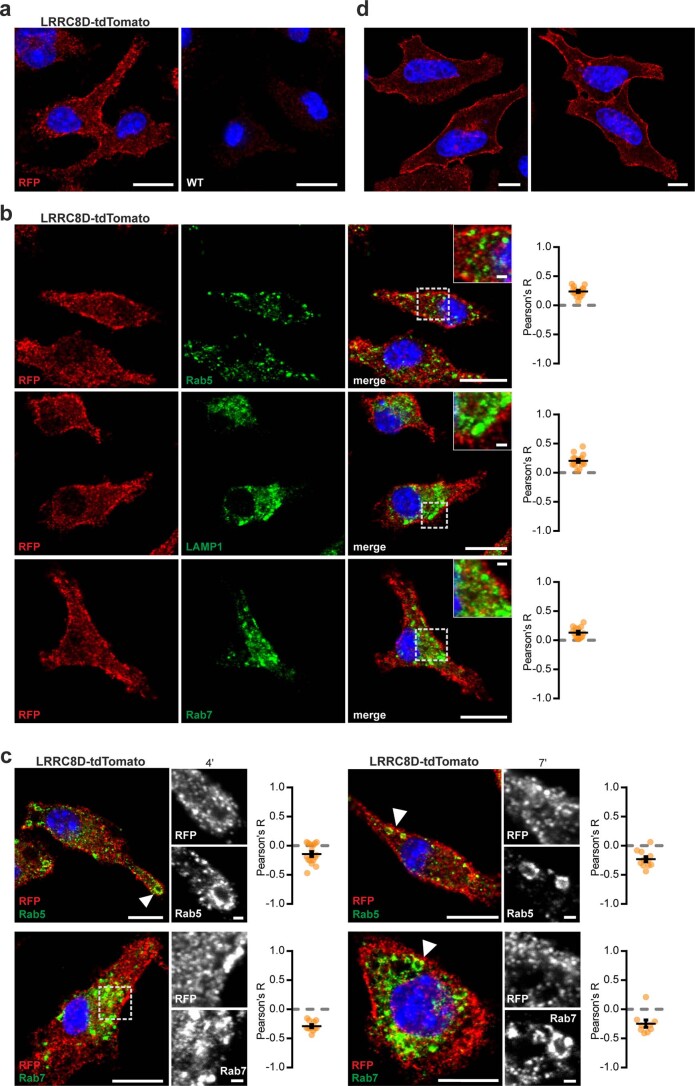
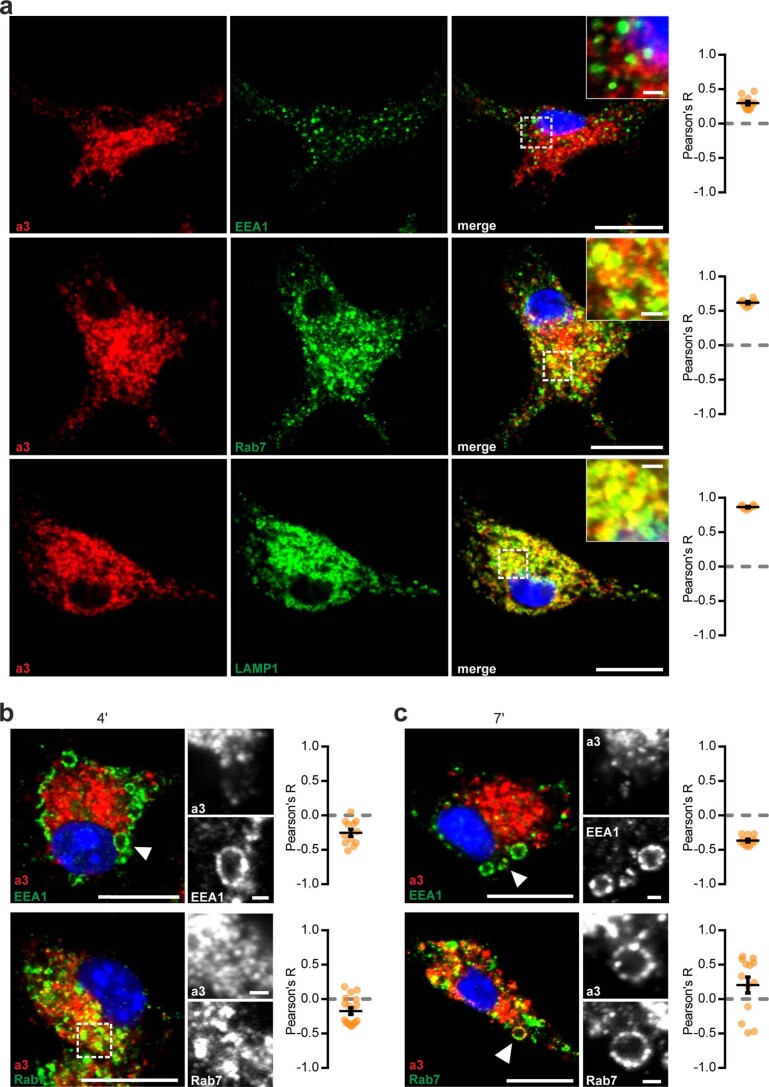
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
