

Nanosurfer assay dissects β-cardiac myosin and cardiac myosin-binding protein C interactions
- PMID: 35591788
- PMCID: PMC9279167
- DOI: 10.1016/j.bpj.2022.05.013
Nanosurfer assay dissects β-cardiac myosin and cardiac myosin-binding protein C interactions
Abstract
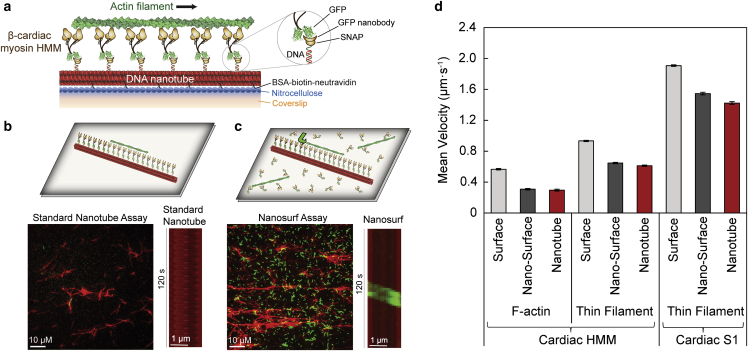
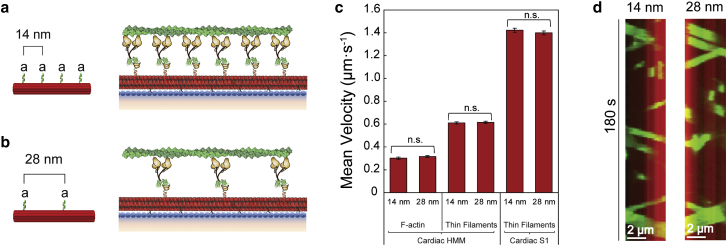
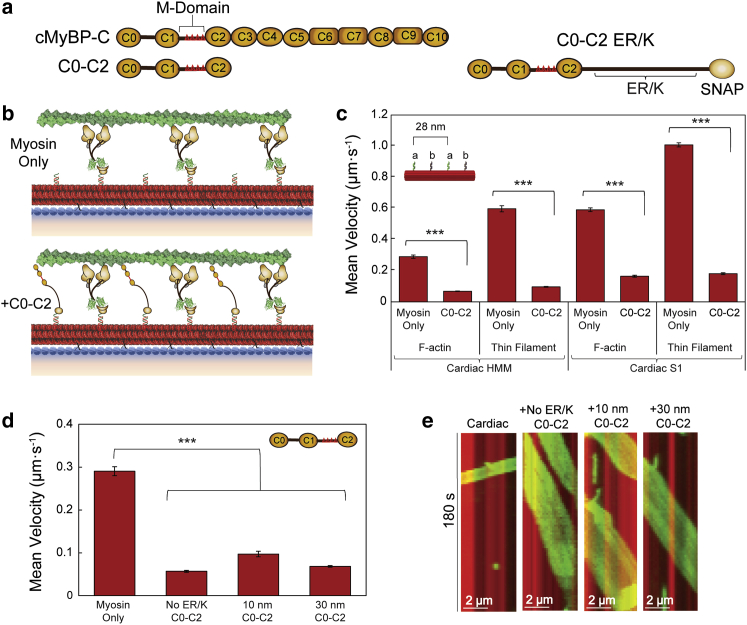
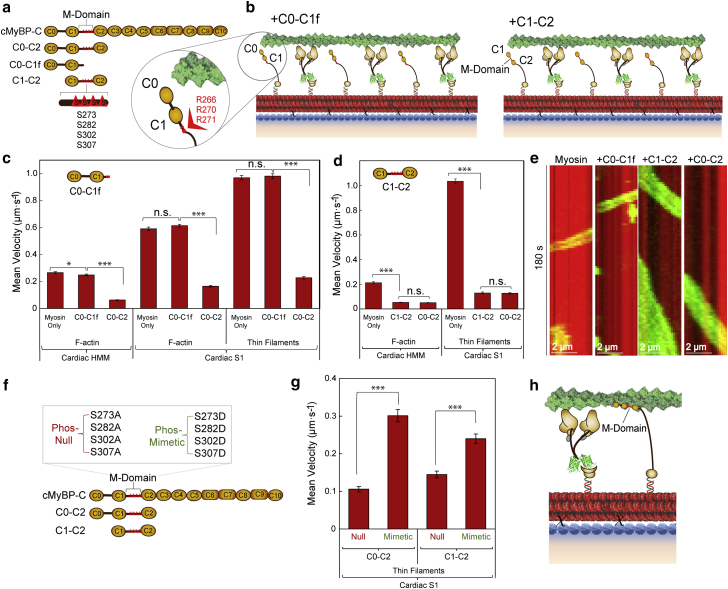
Cardiac myosin-binding protein C (cMyBP-C) modulates cardiac contractility through putative interactions with the myosin S2 tail and/or the thin filament. The relative contribution of these binding-partner interactions to cMyBP-C modulatory function remains unclear. Hence, we developed a "nanosurfer" assay as a model system to interrogate these cMyBP-C binding-partner interactions. Synthetic thick filaments were generated using recombinant human β-cardiac myosin subfragments (HMM or S1) attached to DNA nanotubes, with 14- or 28-nm spacing, corresponding to the 14.3-nm myosin spacing in native thick filaments. The nanosurfer assay consists of DNA nanotubes added to the in vitro motility assay so that myosins on the motility surface effectively deliver thin filaments to the DNA nanotubes, enhancing thin filament gliding probability on the DNA nanotubes. Thin filament velocities on nanotubes with either 14- or 28-nm myosin spacing were no different. We then characterized the effects of cMyBP-C on thin filament motility by alternating HMM and cMyBP-C N-terminal fragments (C0-C2 or C1-C2) on nanotubes every 14 nm. Both C0-C2 and C1-C2 reduced thin filament velocity four- to sixfold relative to HMM alone. Similar inhibition occurred using the myosin S1 construct, which lacks the myosin S2 region proposed to interact with cMyBP-C, suggesting that the cMyBP-C N terminus must interact with other myosin head domains and/or actin to slow thin filament velocity. Thin filament velocity was unaffected by the C0-C1f fragment, which lacks the majority of the M-domain, supporting the importance of this domain for inhibitory interaction(s). A C0-C2 fragment with phospho-mimetic replacement in the M-domain showed markedly less inhibition of thin filament velocity compared with its phospho-null counterpart, highlighting the modulatory role of M-domain phosphorylation on cMyBP-C function. Therefore, the nanosurfer assay provides a platform to precisely manipulate spatially dependent cMyBP-C binding-partner interactions, shedding light on the molecular regulation of β-cardiac myosin contractility.
Copyright © 2022 Biophysical Society. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
