

This is a preprint.
Immediate myeloid depot for SARS-CoV-2 in the human lung
- PMID: 35592107
- PMCID: PMC9119017
- DOI: 10.1101/2022.04.28.489942
Immediate myeloid depot for SARS-CoV-2 in the human lung
Update in
-
Immediate myeloid depot for SARS-CoV-2 in the human lung.Sci Adv. 2024 Aug 2;10(31):eadm8836. doi: 10.1126/sciadv.adm8836. Epub 2024 Jul 31. Sci Adv. 2024. PMID: 39083602 Free PMC article.
Abstract
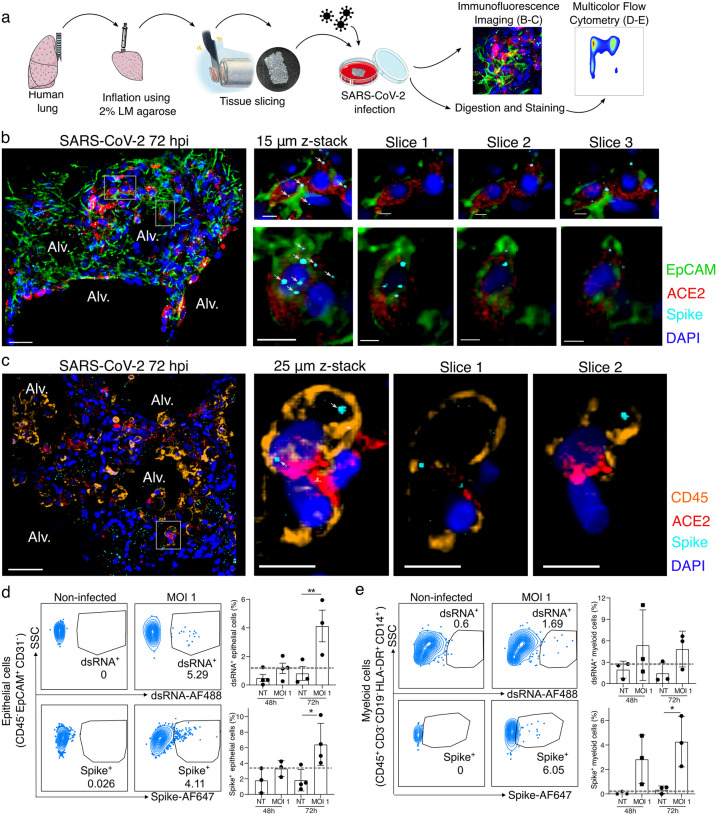
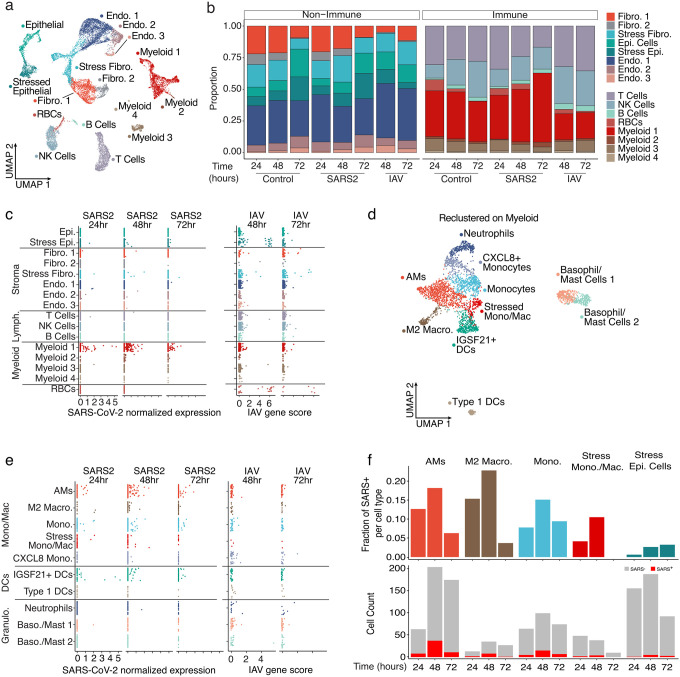
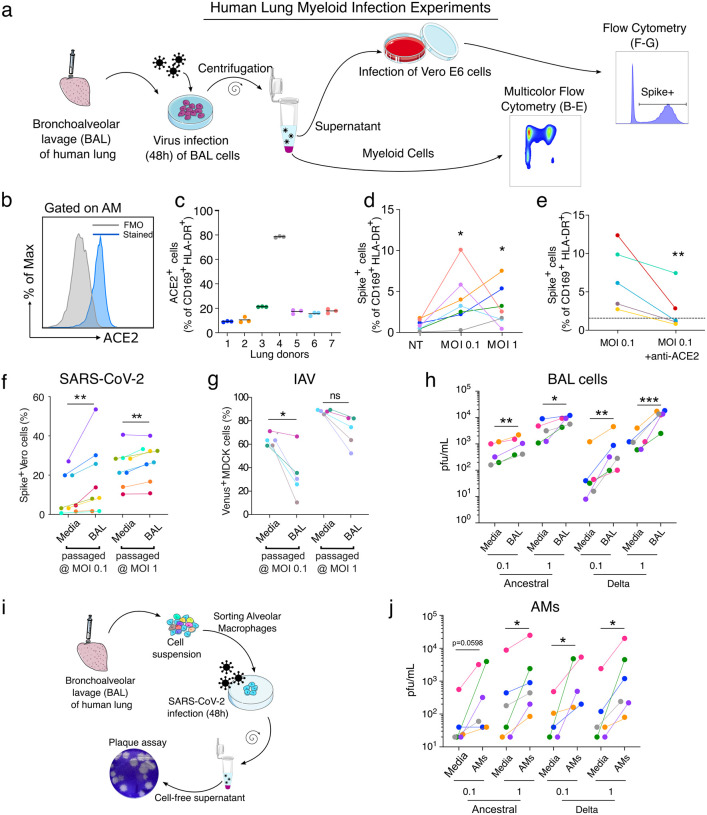
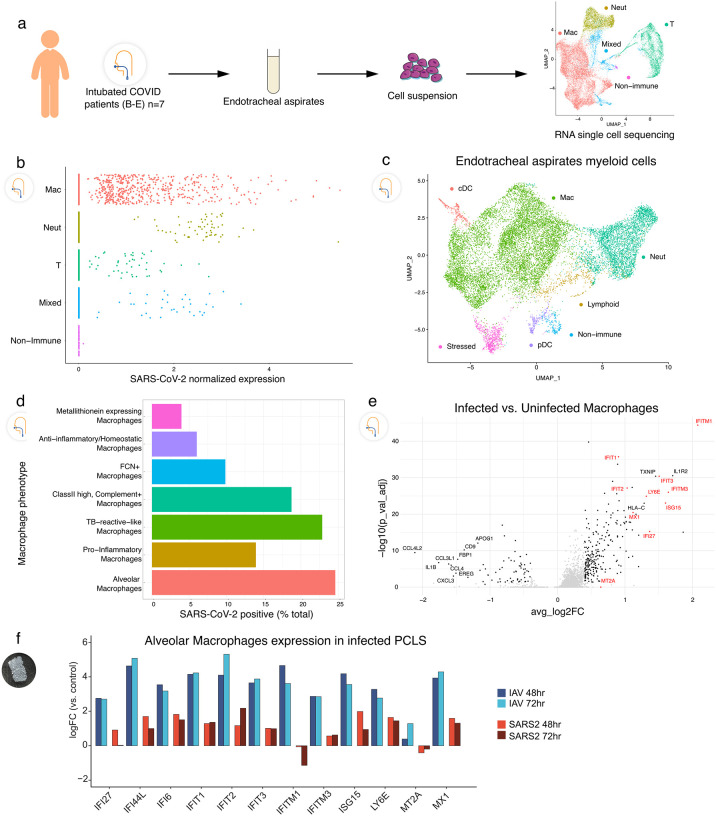
In the severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) pandemic, considerable focus has been placed on a model of viral entry into host epithelial populations, with a separate focus upon the responding immune system dysfunction that exacerbates or causes disease. We developed a precision-cut lung slice model to investigate very early host-viral pathogenesis and found that SARS-CoV-2 had a rapid and specific tropism for myeloid populations in the human lung. Infection of alveolar macrophages was partially dependent upon their expression of ACE2, and the infections were productive for amplifying virus, both findings which were in contrast with their neutralization of another pandemic virus, Influenza A virus (IAV). Compared to IAV, SARS-CoV-2 was extremely poor at inducing interferon-stimulated genes in infected myeloid cells, providing a window of opportunity for modest titers to amplify within these cells. Endotracheal aspirate samples from humans with the acute respiratory distress syndrome (ARDS) from COVID-19 confirmed the lung slice findings, revealing a persistent myeloid depot. In the early phase of SARS-CoV-2 infection, myeloid cells may provide a safe harbor for the virus with minimal immune stimulatory cues being generated, resulting in effective viral colonization and quenching of the immune system.
Conflict of interest statement
Competing interests. The authors declare no competing interests.
Figures
References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous