

Acute bioenergetic insulin sensitivity of skeletal muscle cells: ATP-demand-provoked glycolysis contributes to stimulation of ATP supply
- PMID: 35592612
- PMCID: PMC9112030
- DOI: 10.1016/j.bbrep.2022.101274
Acute bioenergetic insulin sensitivity of skeletal muscle cells: ATP-demand-provoked glycolysis contributes to stimulation of ATP supply
Abstract
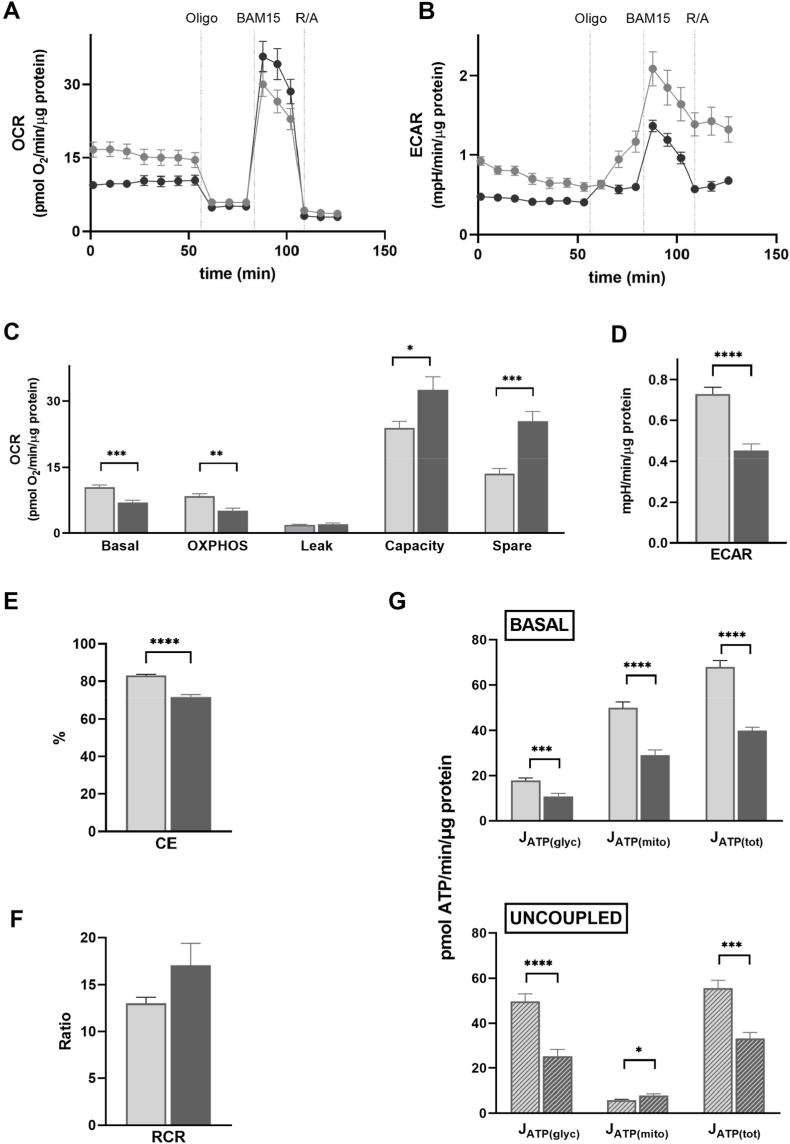
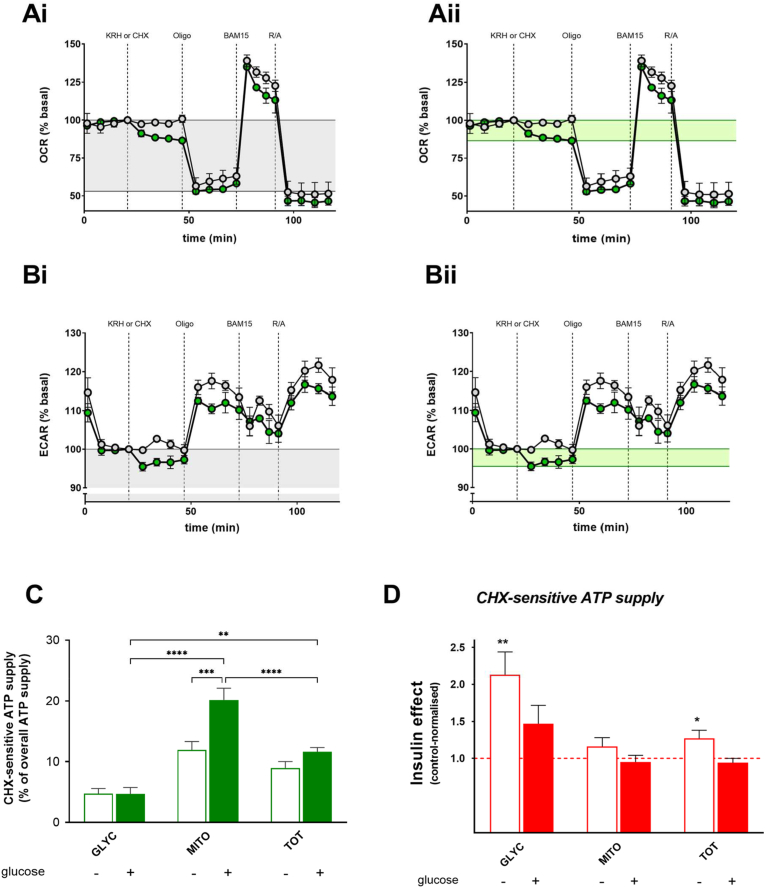
Skeletal muscle takes up glucose in an insulin-sensitive manner and is thus important for the maintenance of blood glucose homeostasis. Insulin resistance during development of type 2 diabetes is associated with decreased ATP synthesis, but the causality of this association is controversial. In this paper, we report real-time oxygen uptake and medium acidification data that we use to quantify acute insulin effects on intracellular ATP supply and ATP demand in rat and human skeletal muscle cells. We demonstrate that insulin increases overall cellular ATP supply by stimulating the rate of glycolytic ATP synthesis. Stimulation is immediate and achieved directly by increased glycolytic capacity, and indirectly by elevated ATP demand from protein synthesis. Raised glycolytic capacity does not result from augmented glucose uptake. Notably, insulin-sensitive glucose uptake is increased synergistically by nitrite. While nitrite has a similar stimulatory effect on glycolytic ATP supply as insulin, it does not amplify insulin stimulation. These data highlight the multifarious nature of acute bioenergetic insulin sensitivity of skeletal muscle cells, and are thus important for the interpretation of changes in energy metabolism that are seen in insulin-resistant muscle.
Keywords: ATP demand; Cellular energy metabolism; Efficiency of mitochondrial ATP synthesis; Oxidative phosphorylation; Skeletal muscle insulin resistance; Type 2 diabetes.
© 2022 The Authors.
Conflict of interest statement
The authors declare no conflict of interest. The sponsors (10.13039/501100000643Daphne Jackson Trust, 10.13039/501100000291Kidney Research UK and the University of Plymouth) had no role in the design, execution, interpretation or writing of the study. It was the decision of the authors only to submit the manuscript for publication.
Figures


References
-
- Thiebaud D., Jacot E., DeFronzo R.A., Maeder E., Jequier E., Felber J.P. The effect of graded doses of insulin on total glucose uptake, glucose oxidation, and glucose storage in man. Diabetes. 1982;31:957–963. - PubMed
-
- Rutter G.A., Pullen T.J., Hodson D.J., Martinez-Sanchez A. Pancreatic β-cell identity, glucose sensing and the control of insulin secretion. Biochem. J. 2015;466:203–218. - PubMed
-
- Garber A.J. Obesity and type 2 diabetes: which patients are at risk? Diabetes Obes. Metabol. 2012;14:399–408. - PubMed
LinkOut - more resources
Full Text Sources
