

Protein lifetimes in aged brains reveal a proteostatic adaptation linking physiological aging to neurodegeneration
- PMID: 35594347
- PMCID: PMC9122331
- DOI: 10.1126/sciadv.abn4437
Protein lifetimes in aged brains reveal a proteostatic adaptation linking physiological aging to neurodegeneration
Abstract
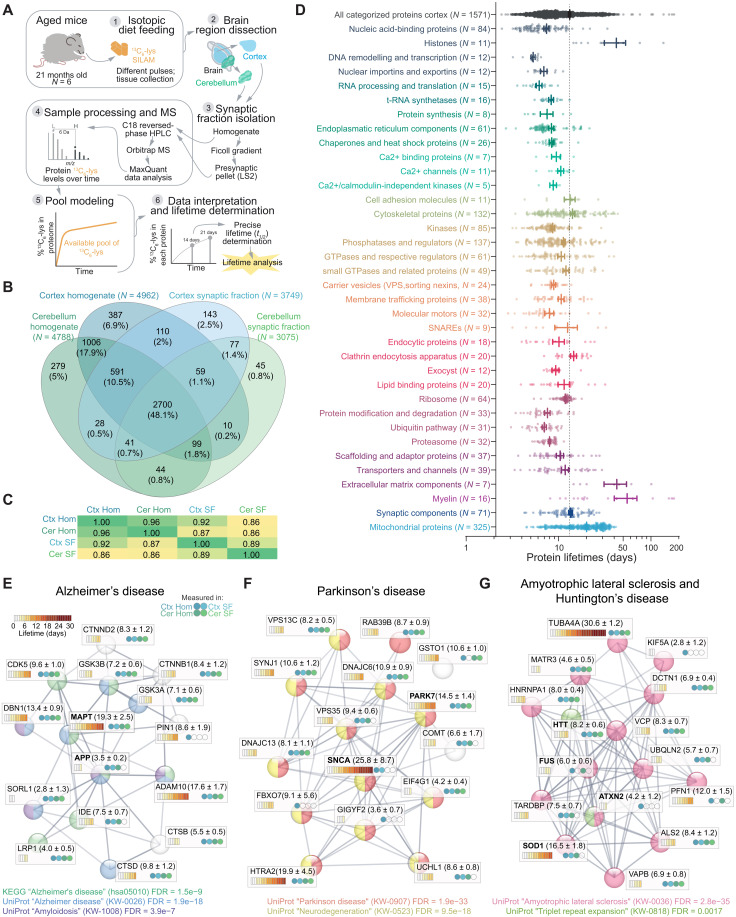
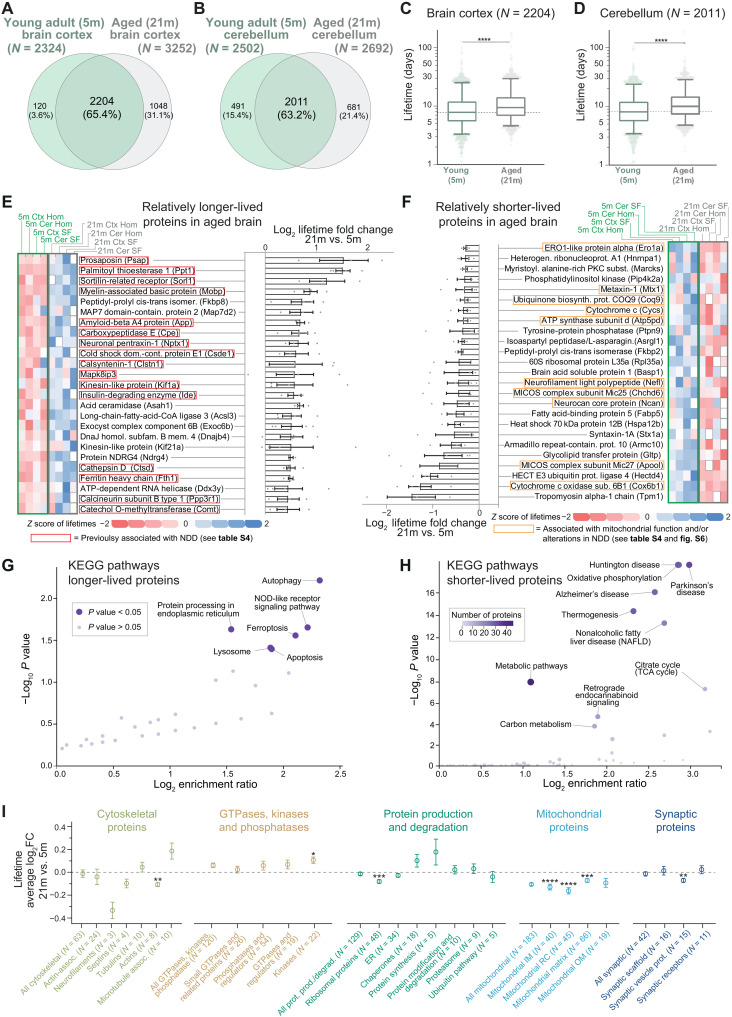
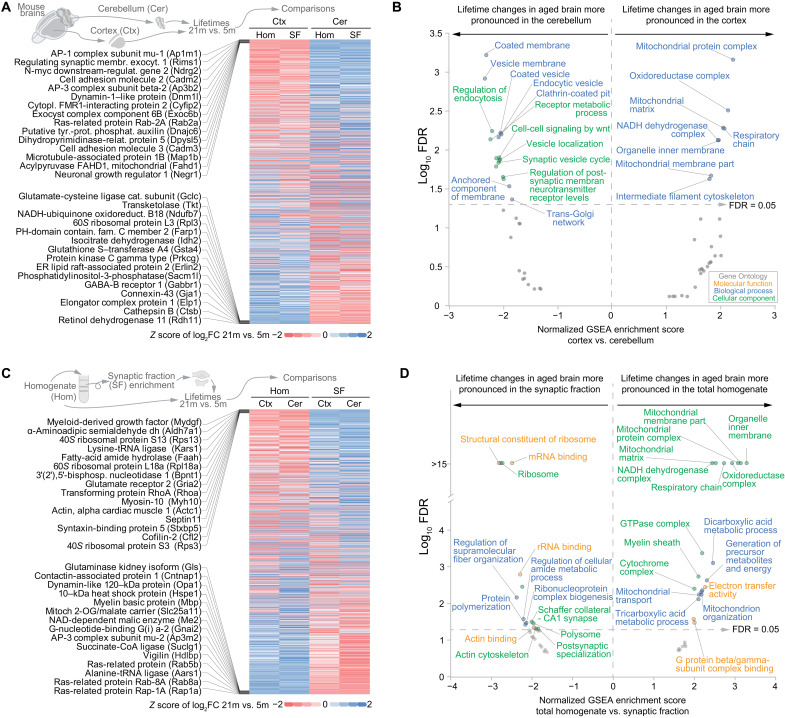
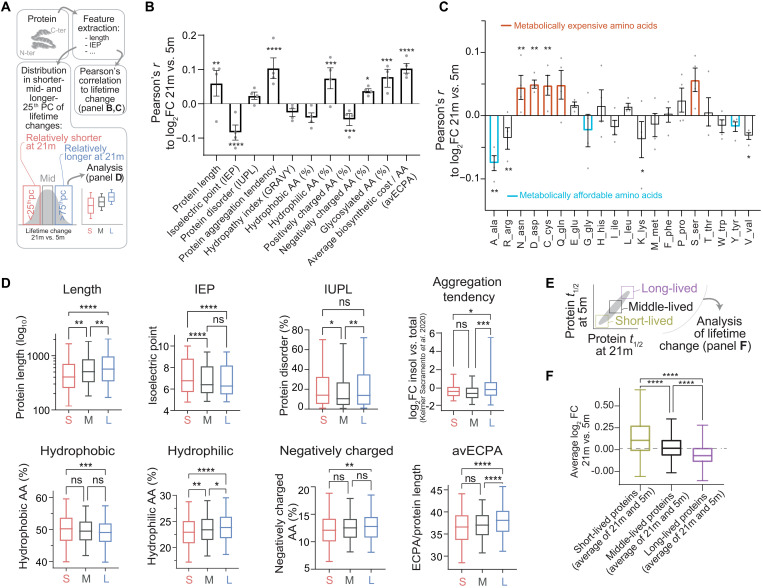
Aging is a prominent risk factor for neurodegenerative disorders (NDDs); however, the molecular mechanisms rendering the aged brain particularly susceptible to neurodegeneration remain unclear. Here, we aim to determine the link between physiological aging and NDDs by exploring protein turnover using metabolic labeling and quantitative pulse-SILAC proteomics. By comparing protein lifetimes between physiologically aged and young adult mice, we found that in aged brains protein lifetimes are increased by ~20% and that aging affects distinct pathways linked to NDDs. Specifically, a set of neuroprotective proteins are longer-lived in aged brains, while some mitochondrial proteins linked to neurodegeneration are shorter-lived. Strikingly, we observed a previously unknown alteration in proteostasis that correlates to parsimonious turnover of proteins with high biosynthetic costs, revealing an overall metabolic adaptation that preludes neurodegeneration. Our findings suggest that future therapeutic paradigms, aimed at addressing these metabolic adaptations, might be able to delay NDD onset.
Figures
References
-
- Hou Y., Dan X., Babbar M., Wei Y., Hasselbalch S. G., Croteau D. L., Bohr V. A., Ageing as a risk factor for neurodegenerative disease. Nat. Rev. Neurol. 15, 565–581 (2019). - PubMed
-
- Kelmer Sacramento E., Kirkpatrick J. M., Mazzetto M., Baumgart M., Bartolome A., Di Sanzo S., Caterino C., Sanguanini M., Papaevgeniou N., Lefaki M., Childs D., Bagnoli S., Terzibasi Tozzini E., Di Fraia D., Romanov N., Sudmant P. H., Huber W., Chondrogianni N., Vendruscolo M., Cellerino A., Ori A., Reduced proteasome activity in the aging brain results in ribosome stoichiometry loss and aggregation. Mol. Syst. Biol. 16, e9596 (2020). - PMC - PubMed
-
- Ward W. F., The relentless effects of the aging process on protein turnover. Biogerontology 1, 195–199 (2000). - PubMed
MeSH terms
LinkOut - more resources
Full Text Sources
Medical
