

Modelling the pyrenoid-based CO2-concentrating mechanism provides insights into its operating principles and a roadmap for its engineering into crops
- PMID: 35596080
- PMCID: PMC9122830
- DOI: 10.1038/s41477-022-01153-7
Modelling the pyrenoid-based CO2-concentrating mechanism provides insights into its operating principles and a roadmap for its engineering into crops
Abstract
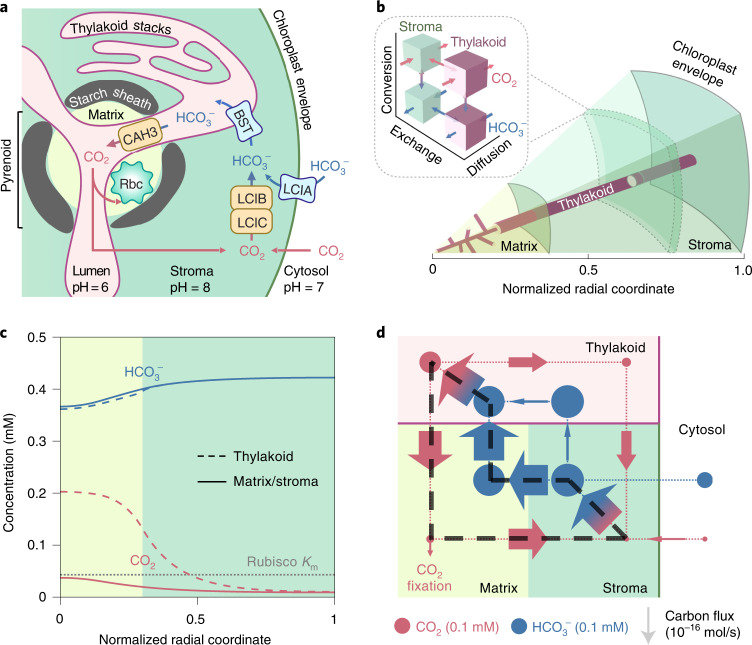
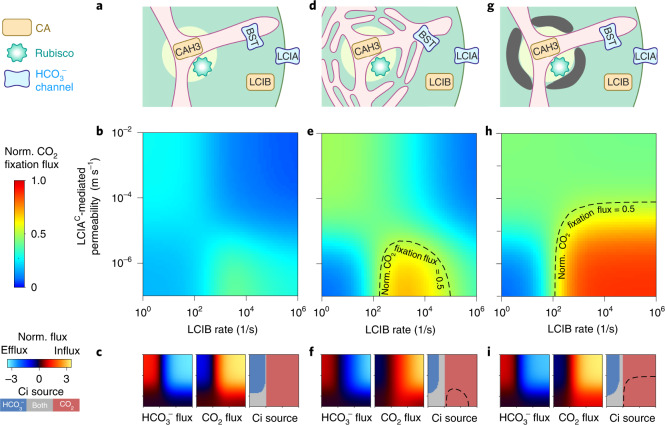
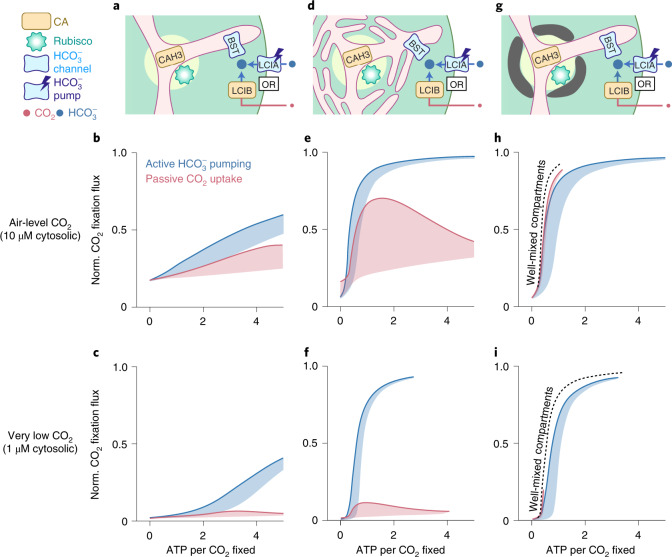
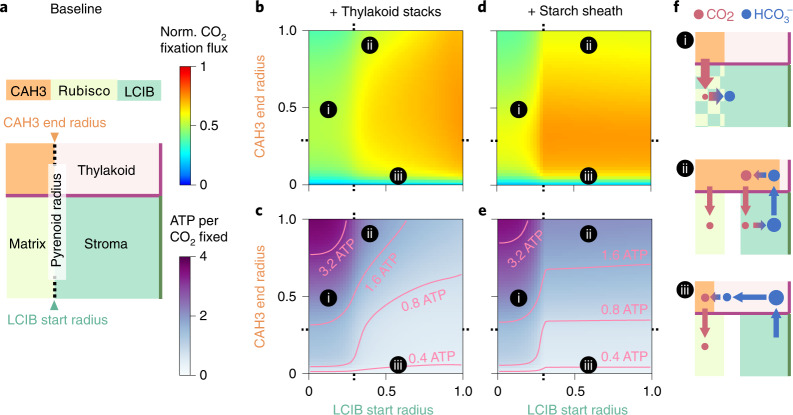
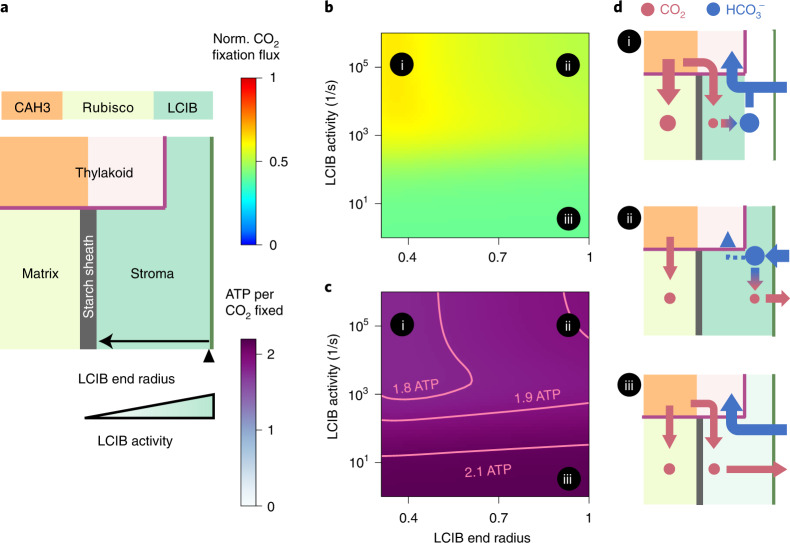
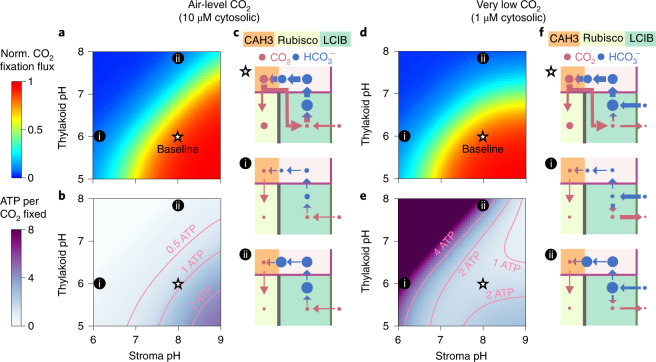
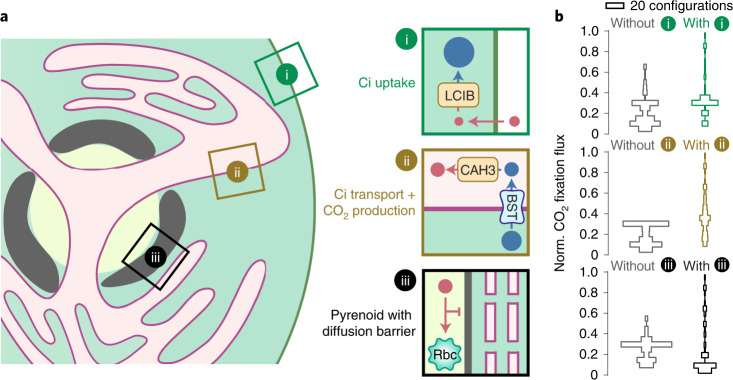
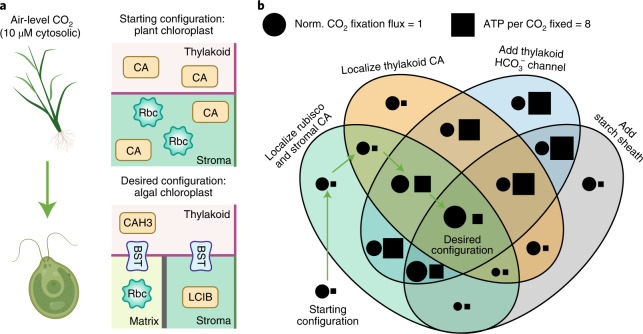
Many eukaryotic photosynthetic organisms enhance their carbon uptake by supplying concentrated CO2 to the CO2-fixing enzyme Rubisco in an organelle called the pyrenoid. Ongoing efforts seek to engineer this pyrenoid-based CO2-concentrating mechanism (PCCM) into crops to increase yields. Here we develop a computational model for a PCCM on the basis of the postulated mechanism in the green alga Chlamydomonas reinhardtii. Our model recapitulates all Chlamydomonas PCCM-deficient mutant phenotypes and yields general biophysical principles underlying the PCCM. We show that an effective and energetically efficient PCCM requires a physical barrier to reduce pyrenoid CO2 leakage, as well as proper enzyme localization to reduce futile cycling between CO2 and HCO3-. Importantly, our model demonstrates the feasibility of a purely passive CO2 uptake strategy at air-level CO2, while active HCO3- uptake proves advantageous at lower CO2 levels. We propose a four-step engineering path to increase the rate of CO2 fixation in the plant chloroplast up to threefold at a theoretical cost of only 1.3 ATP per CO2 fixed, thereby offering a framework to guide the engineering of a PCCM into land plants.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
