

A hexasome is the preferred substrate for the INO80 chromatin remodeling complex, allowing versatility of function
- PMID: 35597239
- PMCID: PMC9351570
- DOI: 10.1016/j.molcel.2022.04.026
A hexasome is the preferred substrate for the INO80 chromatin remodeling complex, allowing versatility of function
Abstract
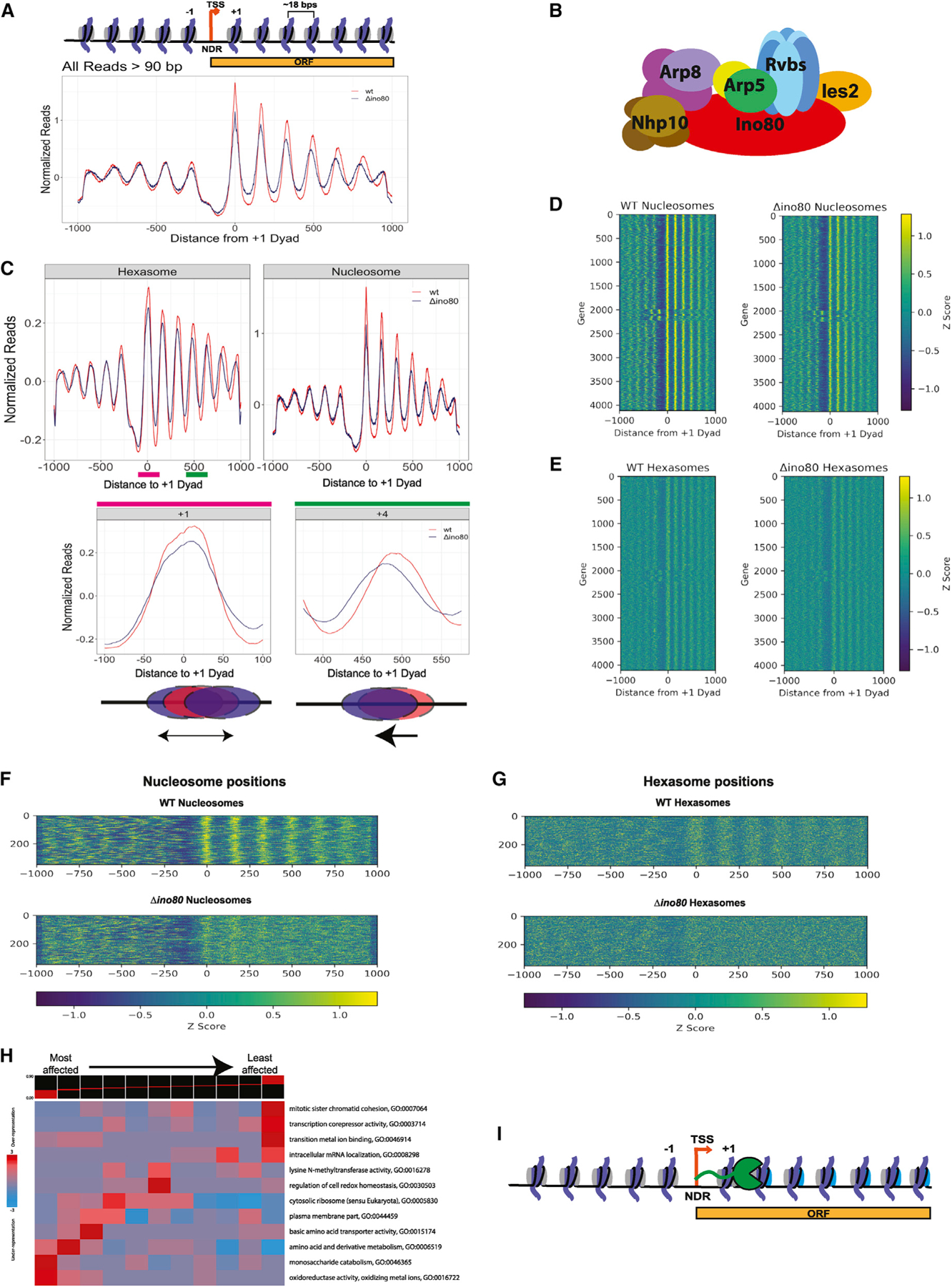
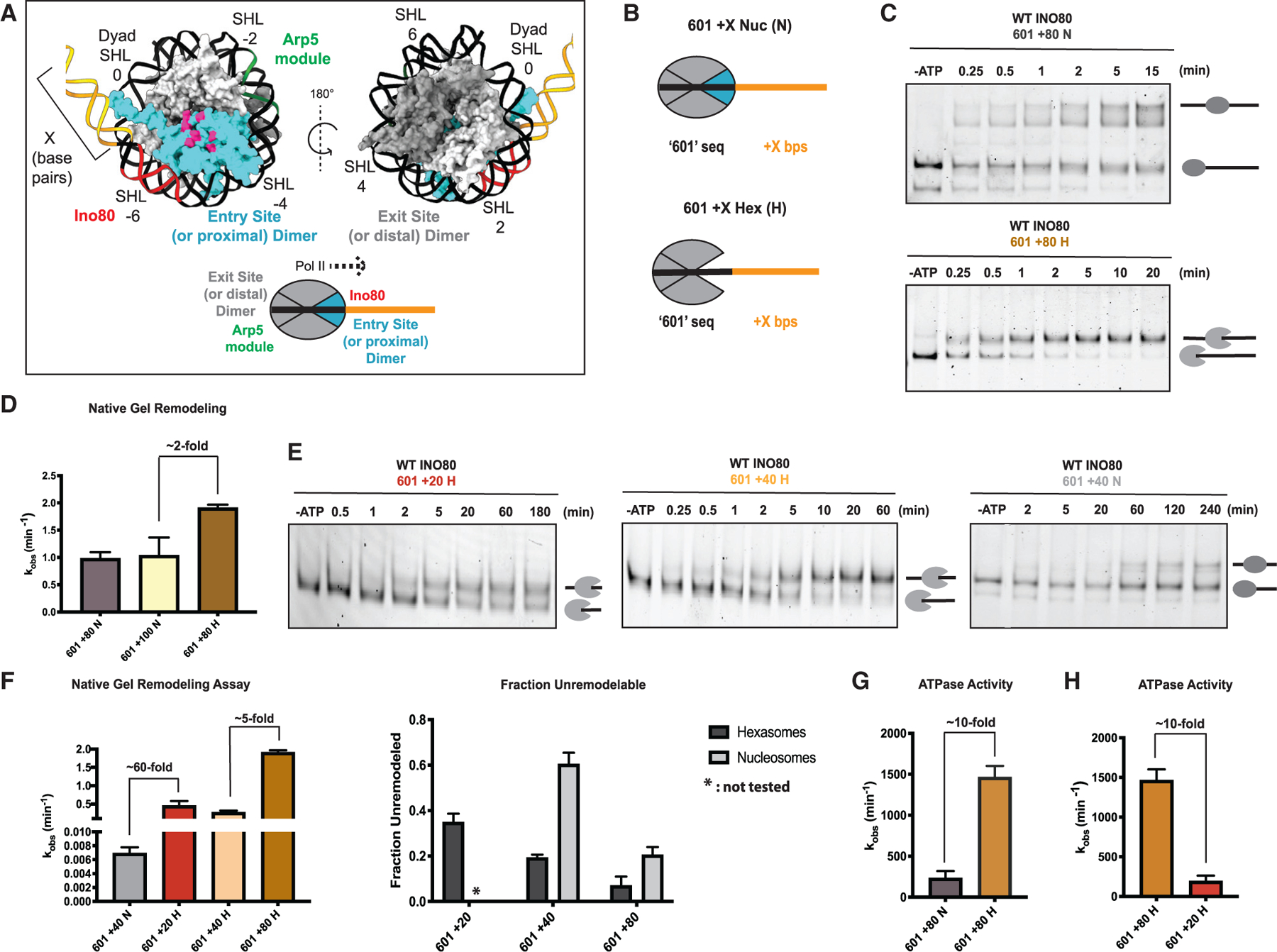
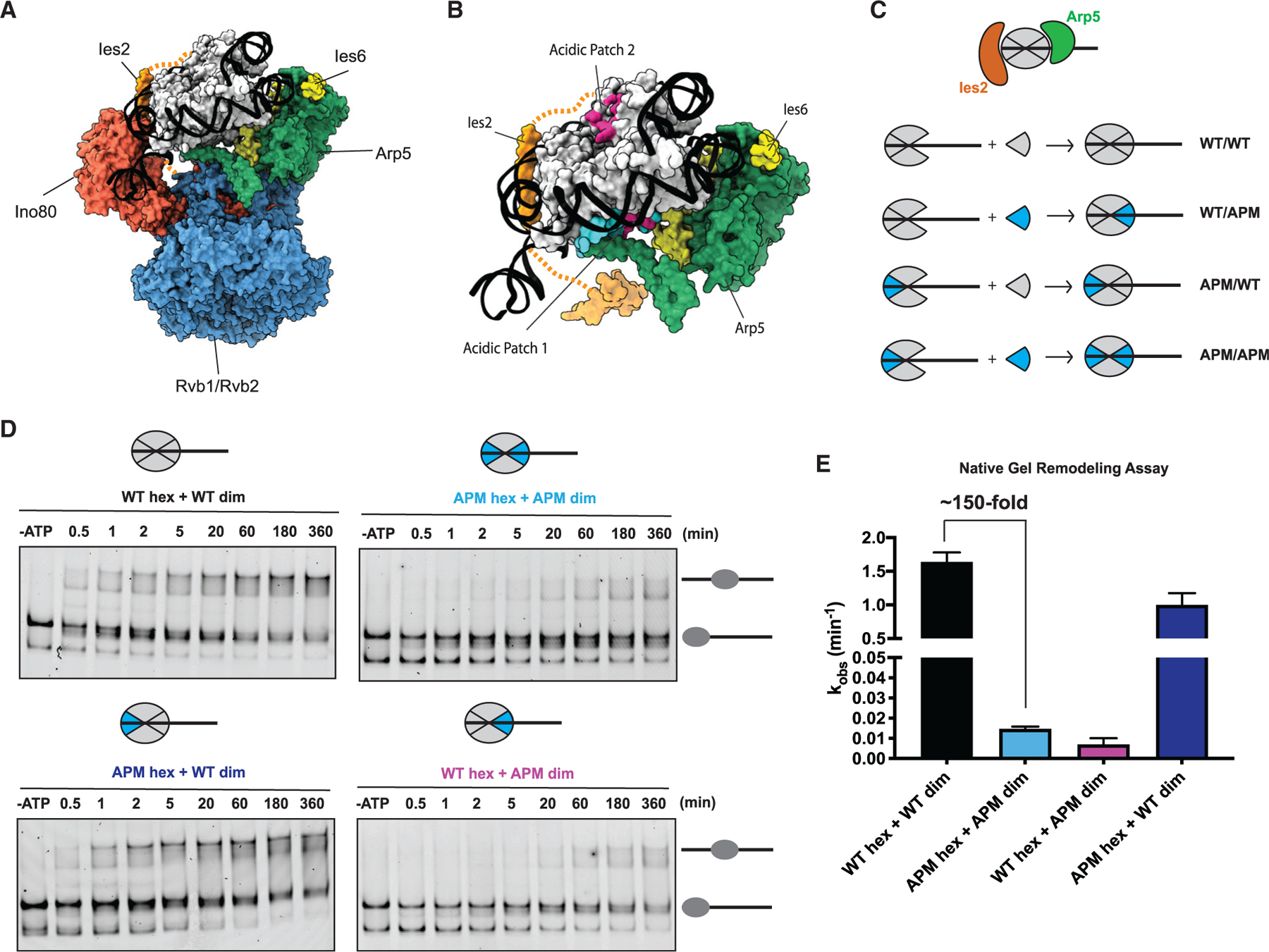
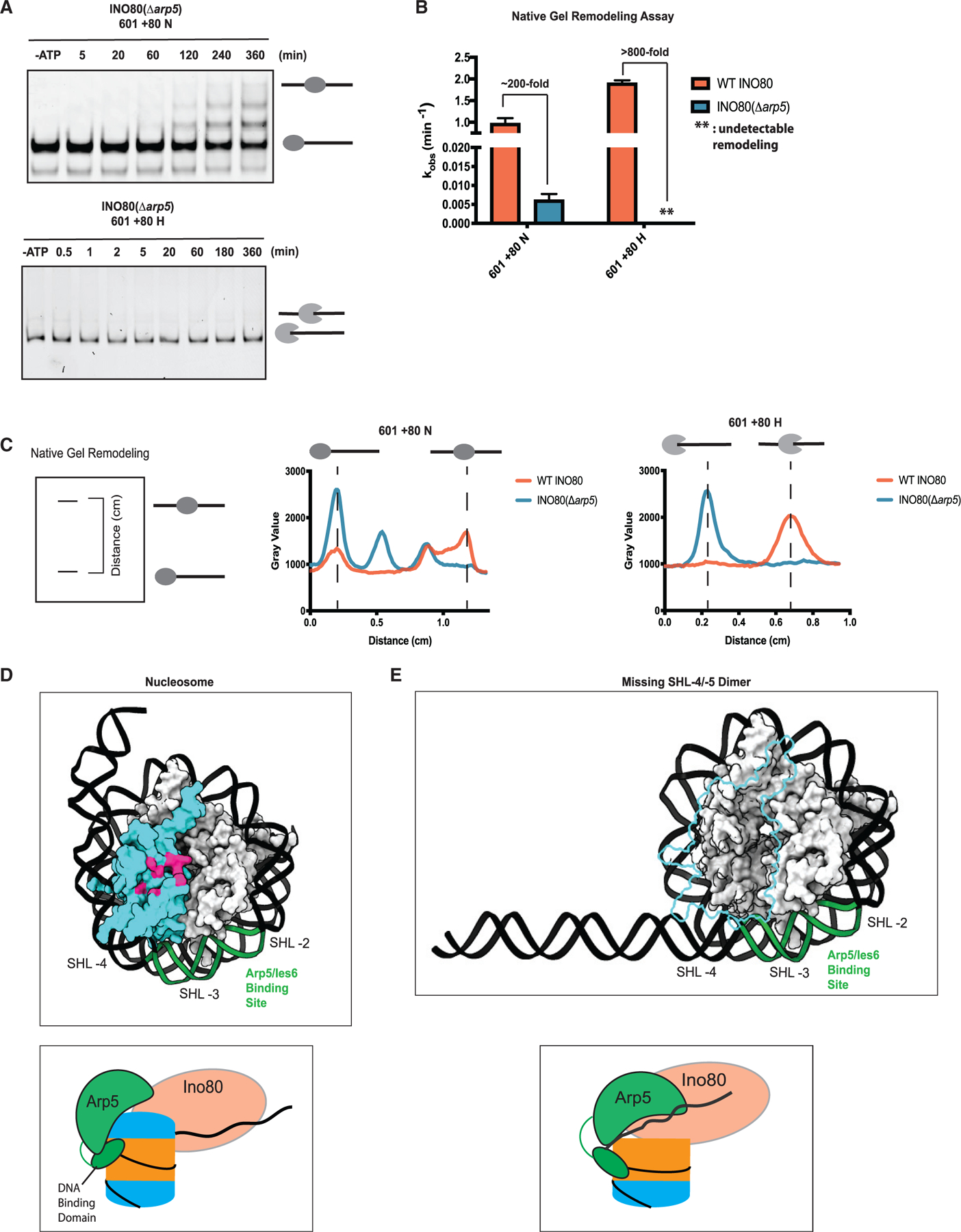
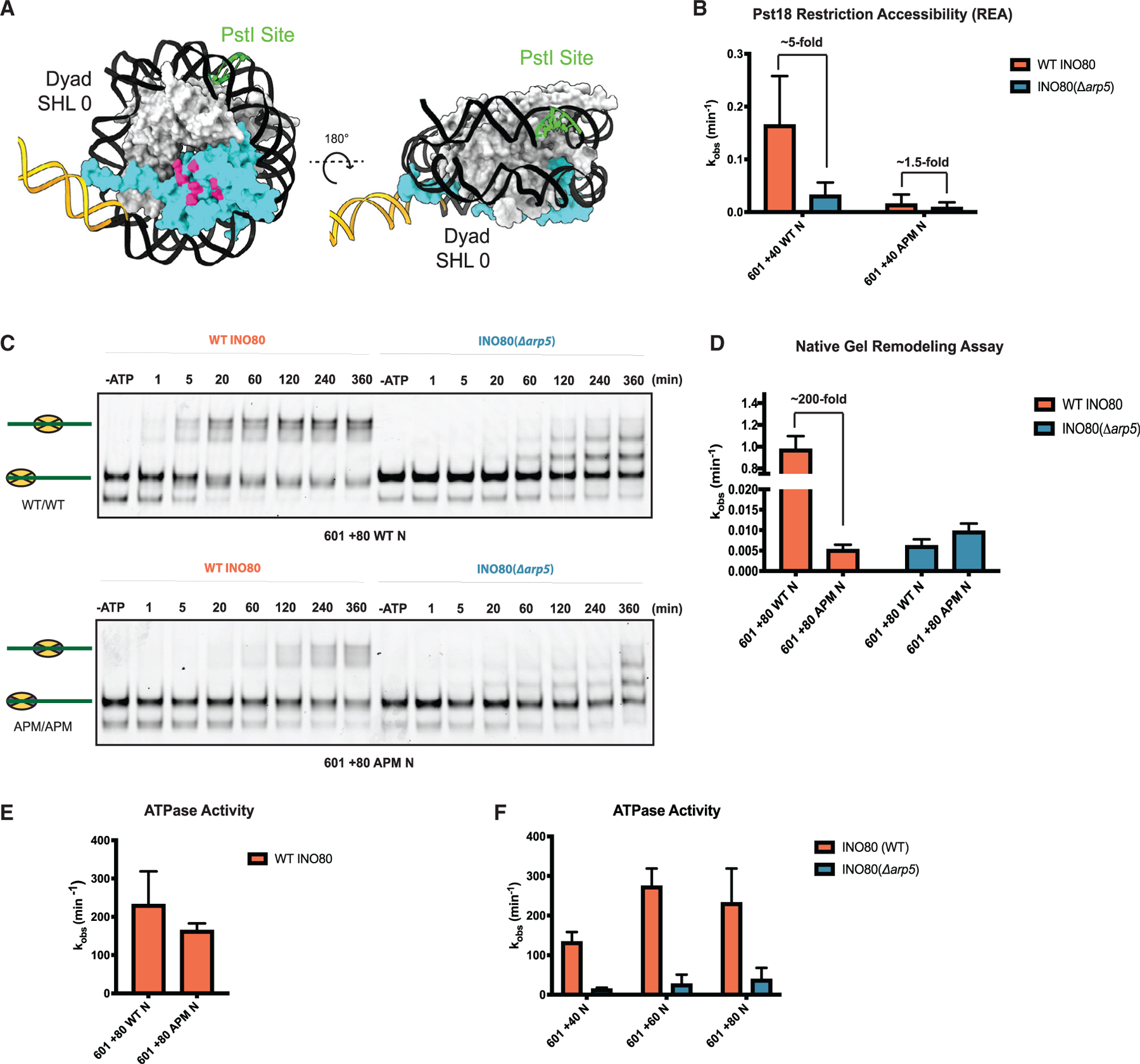
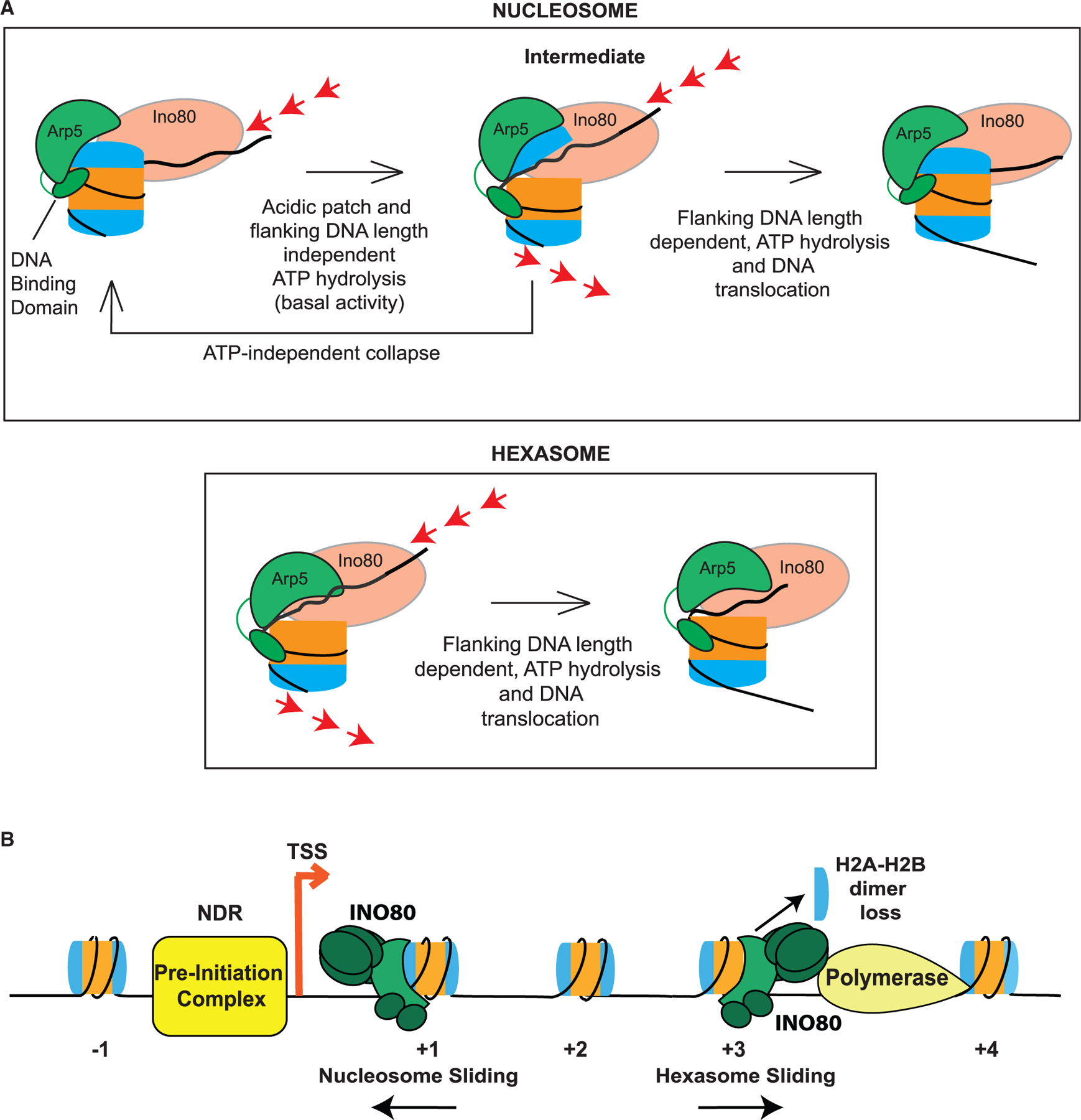
The critical role of the INO80 chromatin remodeling complex in transcription is commonly attributed to its nucleosome sliding activity. Here, we have found that INO80 prefers to mobilize hexasomes over nucleosomes. INO80's preference for hexasomes reaches up to ∼60 fold when flanking DNA overhangs approach ∼18-bp linkers in yeast gene bodies. Correspondingly, deletion of INO80 significantly affects the positions of hexasome-sized particles within yeast genes in vivo. Our results raise the possibility that INO80 promotes nucleosome sliding by dislodging an H2A-H2B dimer, thereby making a nucleosome transiently resemble a hexasome. We propose that this mechanism allows INO80 to rapidly mobilize nucleosomes at promoters and hexasomes within gene bodies. Rapid repositioning of hexasomes that are generated in the wake of transcription may mitigate spurious transcription. More generally, such versatility may explain how INO80 regulates chromatin architecture during the diverse processes of transcription, replication, and repair.
Keywords: INO80; chromatin remodeling; hexasome.
Copyright © 2022 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests Geeta Narlikar is on the Molecular Cell advisory board.
Figures
References
-
- Belotserkovskaya R, Oh S, Bondarenko VA, Orphanides G, Studitsky VM, and Reinberg D (2003). FACT facilitates transcription-dependent nucleosome alteration. Science 301, 1090–1093. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
