

An orally active plant Rubisco-derived peptide increases neuronal leptin responsiveness
- PMID: 35597815
- PMCID: PMC9124197
- DOI: 10.1038/s41598-022-12595-6
An orally active plant Rubisco-derived peptide increases neuronal leptin responsiveness
Abstract
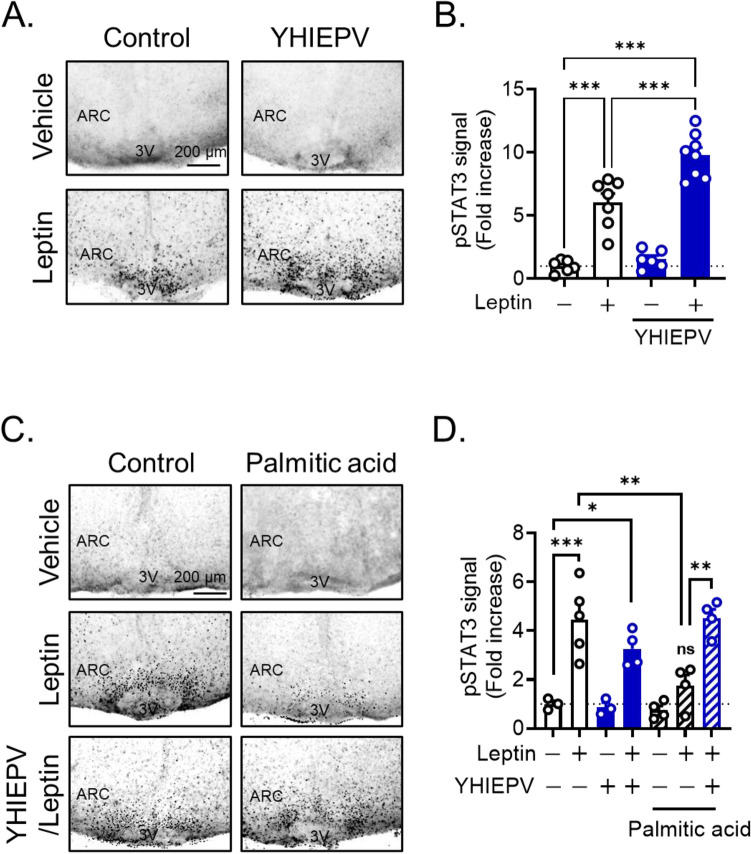
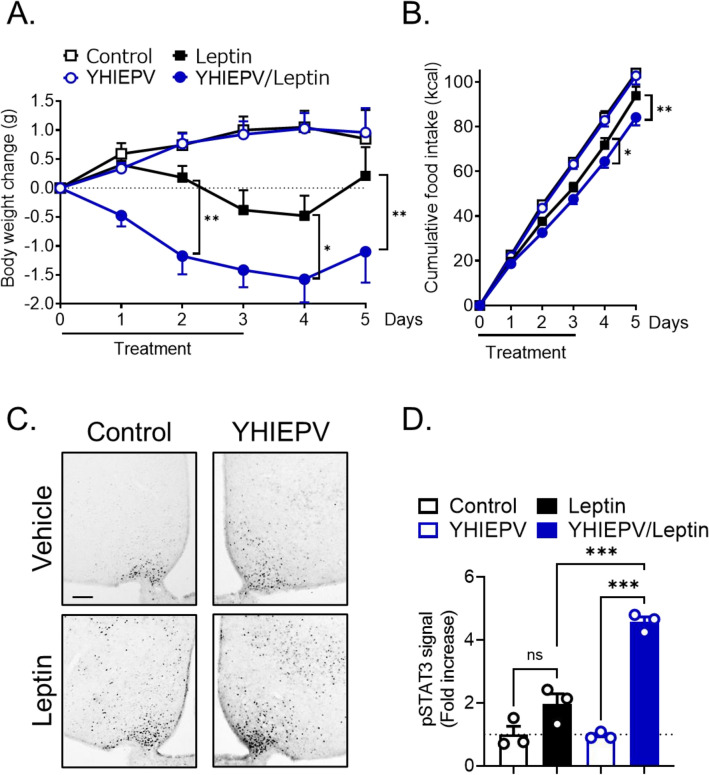
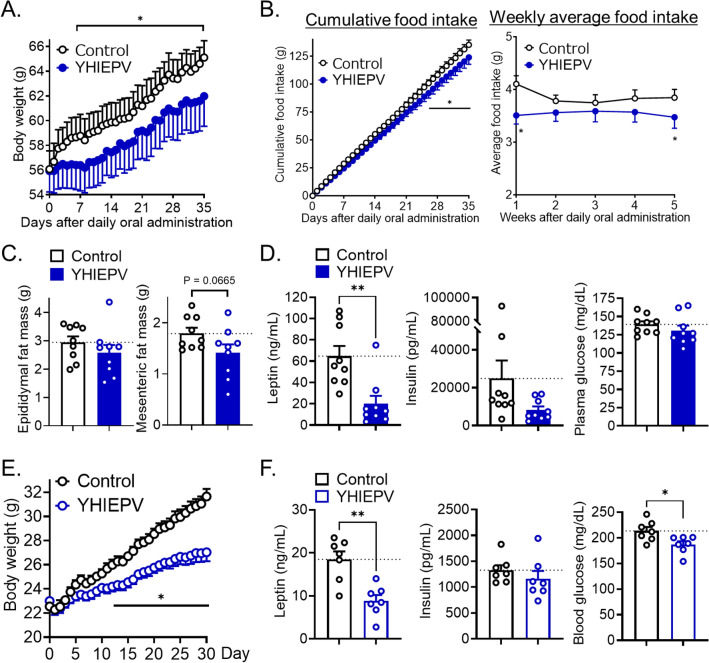
Nutrient excess, such as the intake of a high-fat diet, reduces hypothalamic responses to exogenously administered leptin and induces dietary obesity; however, orally active components that attenuate neural leptin dysregulation have yet to be identified. We herein demonstrated that YHIEPV, derived from the pepsin-pancreatin digestion of the green leaf protein Rubisco, increased the leptin-induced phosphorylation of STAT3 in ex vivo hypothalamic slice cultures. We also showed that YHIEPV mitigated palmitic acid-induced decreases in leptin responsiveness. Furthermore, orally administered YHIEPV promoted leptin-induced reductions in body weight and food intake in obese mice. In addition, dietary-induced body weight gain was significantly less in mice orally or centrally administered YHIEPV daily than in saline-control mice. Cellular leptin sensitivity and the levels of proinflammatory-related factors, such as IL1β and Socs-3, in the hypothalamus of obese mice were also restored by YHIEPV. YHIEPV blocked cellular leptin resistance induced by forskolin, which activates Epac-Rap1 signaling, and reduced the level of the GTP-bound active form of Rap1 in the brains of obese mice. Collectively, the present results demonstrated that the orally active peptide YHIEPV derived from a major green leaf protein increased neural leptin responsiveness and reduced body weight gain in mice with dietary obesity.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures

Similar articles
-
Gut-derived GIP activates central Rap1 to impair neural leptin sensitivity during overnutrition.J Clin Invest. 2019 Aug 12;129(9):3786-3791. doi: 10.1172/JCI126107. eCollection 2019 Aug 12. J Clin Invest. 2019. PMID: 31403469 Free PMC article.
-
Sulforaphane improves leptin responsiveness in high-fat high-sucrose diet-fed obese mice.Eur J Pharmacol. 2018 Sep 15;835:108-114. doi: 10.1016/j.ejphar.2018.07.050. Epub 2018 Jul 29. Eur J Pharmacol. 2018. PMID: 30063917 Free PMC article.
-
High-fat diet-induced changes in body mass and hypothalamic gene expression in wild-type and leptin-deficient mice.Endocrine. 2008 Apr;33(2):176-88. doi: 10.1007/s12020-008-9070-1. Epub 2008 May 16. Endocrine. 2008. PMID: 18483882
-
The evidence of metabolic-improving effect of metformin in Ay/a mice with genetically-induced melanocortin obesity and the contribution of hypothalamic mechanisms to this effect.PLoS One. 2019 Mar 14;14(3):e0213779. doi: 10.1371/journal.pone.0213779. eCollection 2019. PLoS One. 2019. PMID: 30870482 Free PMC article.
-
Activation of the hypothalamic arcuate nucleus predicts the anorectic actions of ciliary neurotrophic factor and leptin in intact and gold thioglucose-lesioned mice.J Neuroendocrinol. 2003 Jul;15(7):649-60. doi: 10.1046/j.1365-2826.2003.01043.x. J Neuroendocrinol. 2003. PMID: 12787049
Cited by
-
Lard intake results in better hypothalamic leptin responsiveness than beef tallow intake during overnutrition.PLoS One. 2025 Jul 21;20(7):e0326847. doi: 10.1371/journal.pone.0326847. eCollection 2025. PLoS One. 2025. PMID: 40690443 Free PMC article.
-
Plant hormone jasmonic acid reduces anxiety behavior in mice.Sci Rep. 2025 Apr 3;15(1):11424. doi: 10.1038/s41598-025-95689-1. Sci Rep. 2025. PMID: 40181068 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
