

The Taspase1/Myosin1f-axis regulates filopodia dynamics
- PMID: 35601920
- PMCID: PMC9121324
- DOI: 10.1016/j.isci.2022.104355
The Taspase1/Myosin1f-axis regulates filopodia dynamics
Abstract
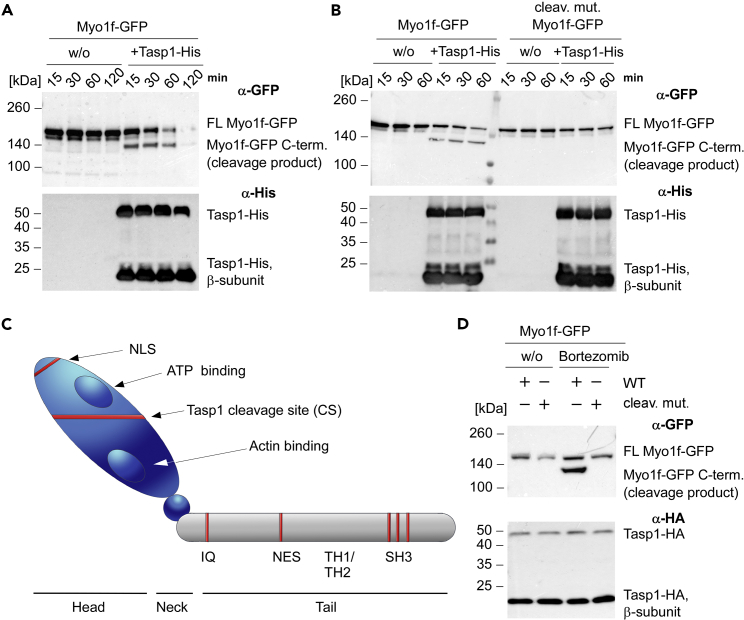
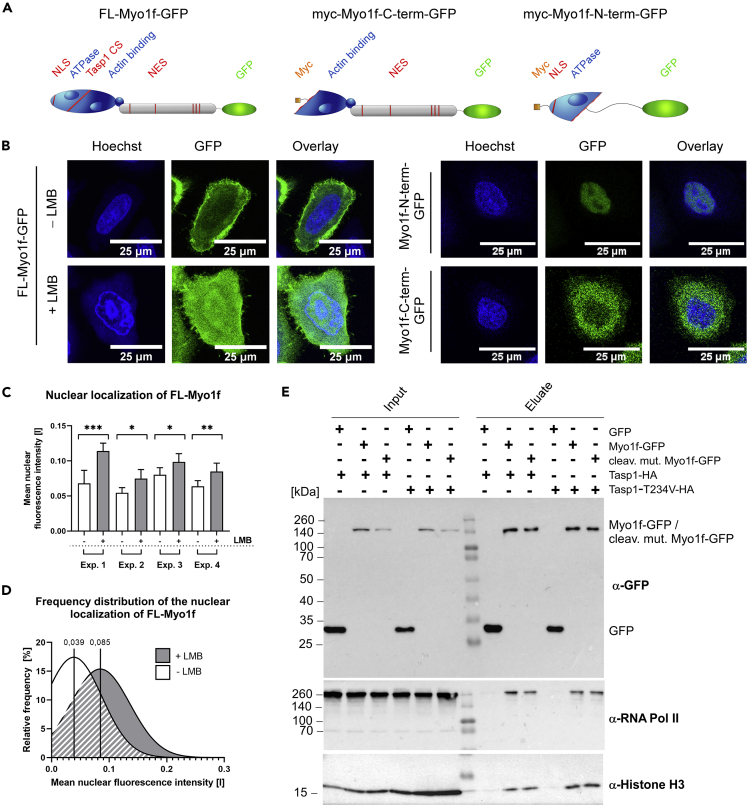
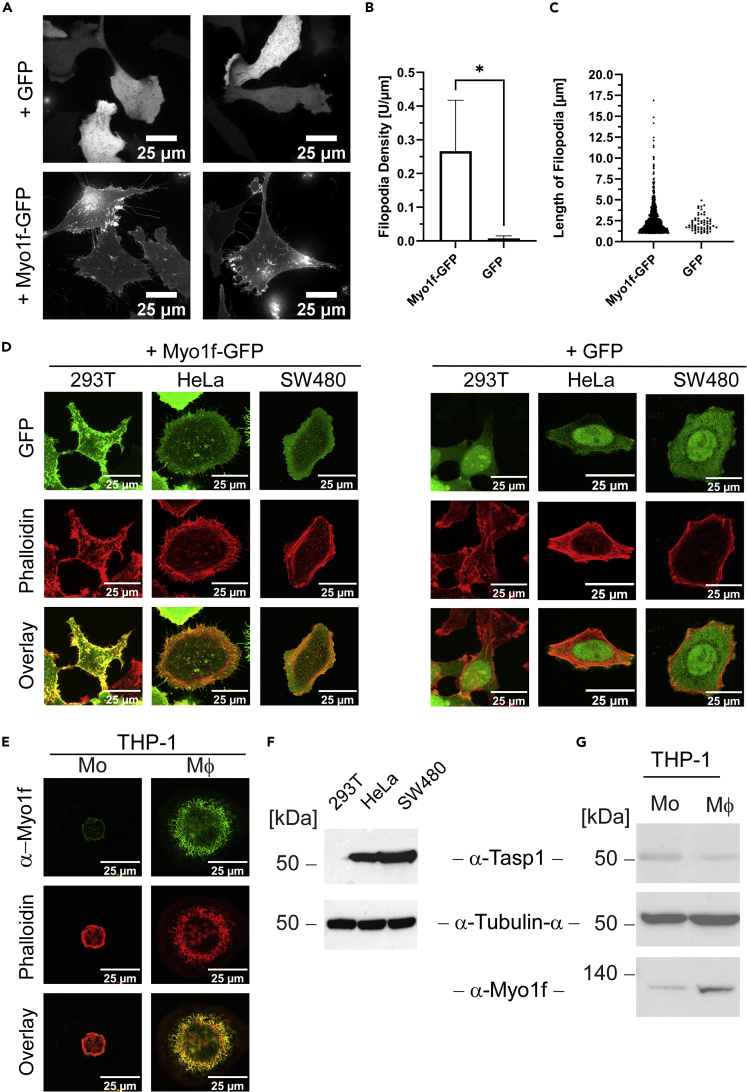
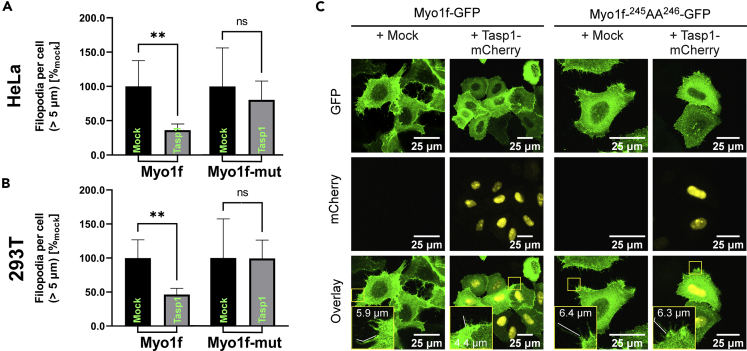
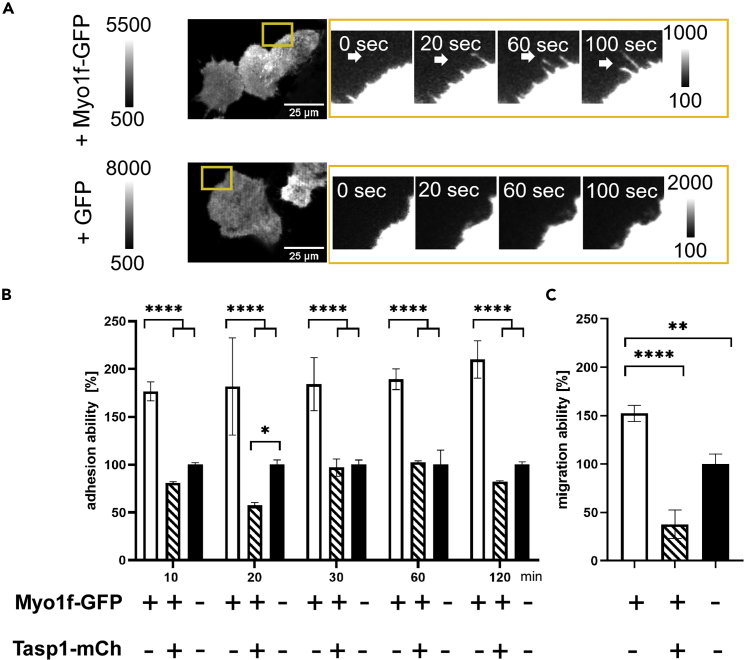
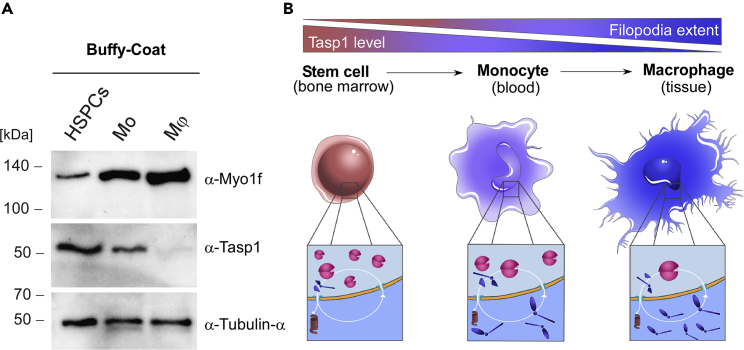
The unique threonine protease Tasp1 impacts not only ordered development and cell proliferation but also pathologies. However, its substrates and the underlying molecular mechanisms remain poorly understood. We demonstrate that the unconventional Myo1f is a Tasp1 substrate and unravel the physiological relevance of this proteolysis. We classify Myo1f as a nucleo-cytoplasmic shuttle protein, allowing its unhindered processing by nuclear Tasp1 and an association with chromatin. Moreover, we show that Myo1f induces filopodia resulting in increased cellular adhesion and migration. Importantly, filopodia formation was antagonized by Tasp1-mediated proteolysis, supported by an inverse correlation between Myo1f concentration and Tasp1 expression level. The Tasp1/Myo1f-axis might be relevant in human hematopoiesis as reduced Tasp1 expression coincided with increased Myo1f concentrations and filopodia in macrophages compared to monocytes and vice versa. In sum, we discovered Tasp1-mediated proteolysis of Myo1f as a mechanism to fine-tune filopodia formation, inter alia relevant for cells of the immune system.
Keywords: Biological sciences; Cell biology; Immunology.
© 2022 The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures

References
-
- Adams J., Palombella V.J., Sausville E.A., Johnson J., Destree A., Lazarus D.D., Maas J., Pien C.S., Prakash S., Elliott P.J. Proteasome inhibitors: a novel class of potent and effective antitumor agents. Cancer Res. 1999;59:2615–2622. - PubMed
-
- Balkin D.M., Poranki M., Forester C.M., Dorsey M.J., Slavotinek A., Pomerantz J.H. TASP1 mutation in a female with craniofacial anomalies, anterior segment dysgenesis, congenital immunodeficiency and macrocytic anemia. Mol. Genet. Genomic Med. 2019;7:e818. doi: 10.1002/mgg3.818. - DOI - PMC - PubMed
LinkOut - more resources
Full Text Sources
