

Changing paradigms in oncology: Toward noncytotoxic treatments for advanced gliomas
- PMID: 35603902
- PMCID: PMC9474618
- DOI: 10.1002/ijc.34131
Changing paradigms in oncology: Toward noncytotoxic treatments for advanced gliomas
Abstract
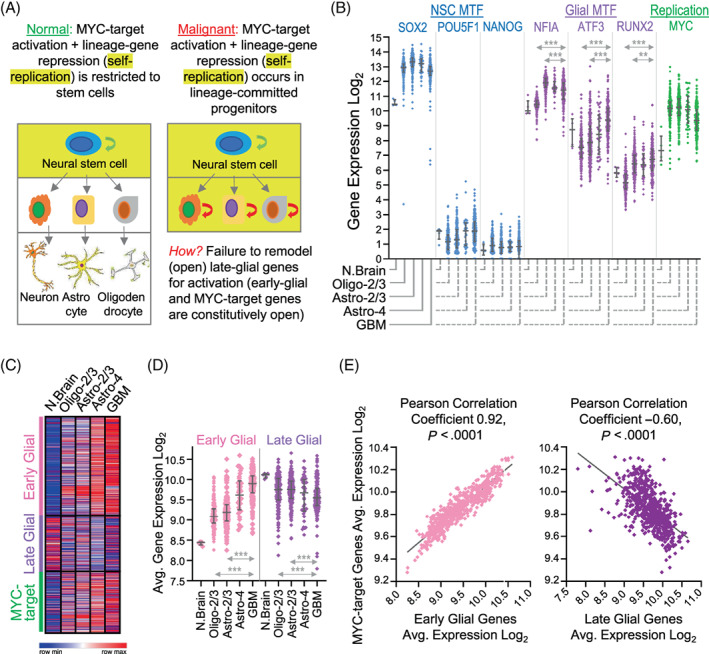
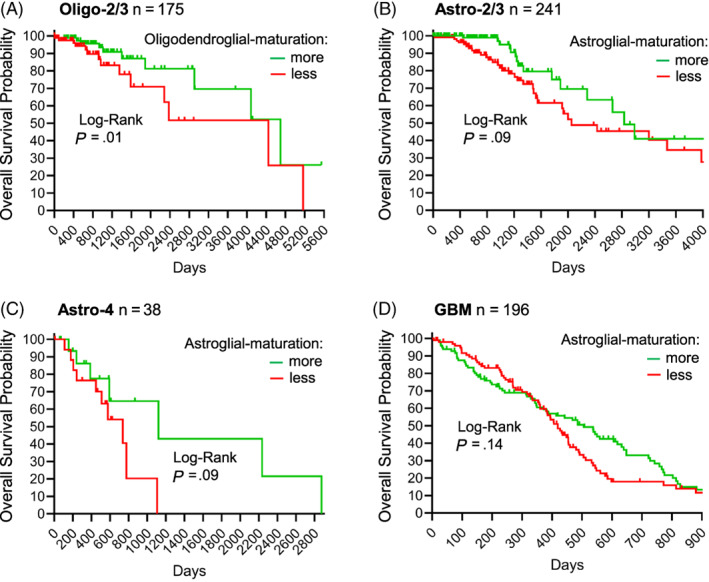
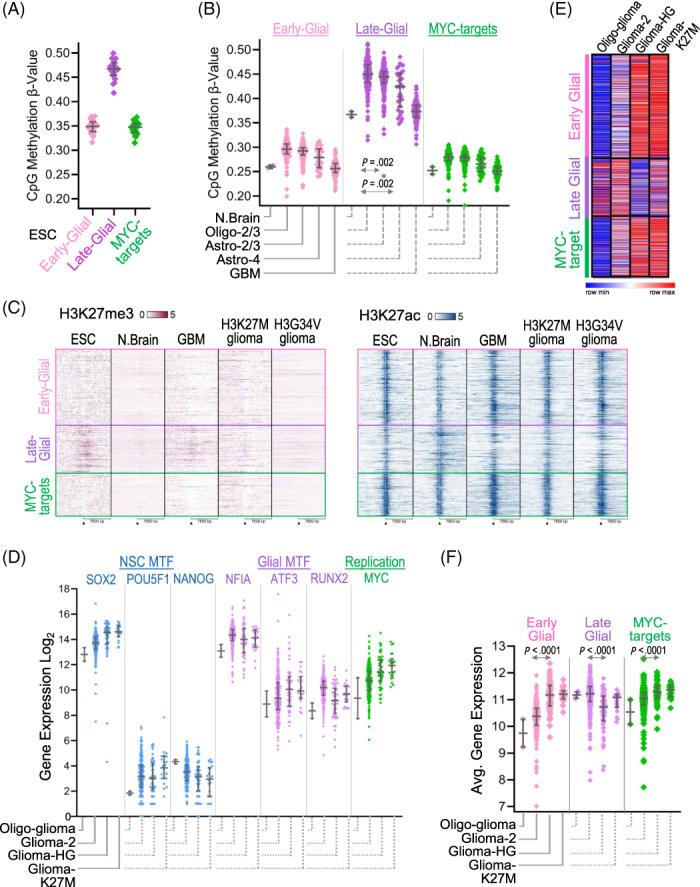
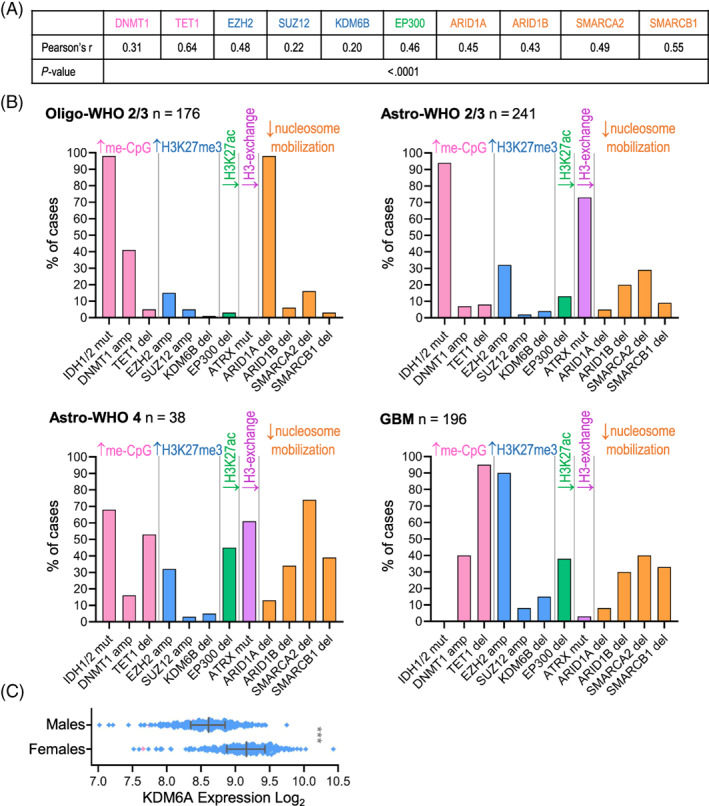
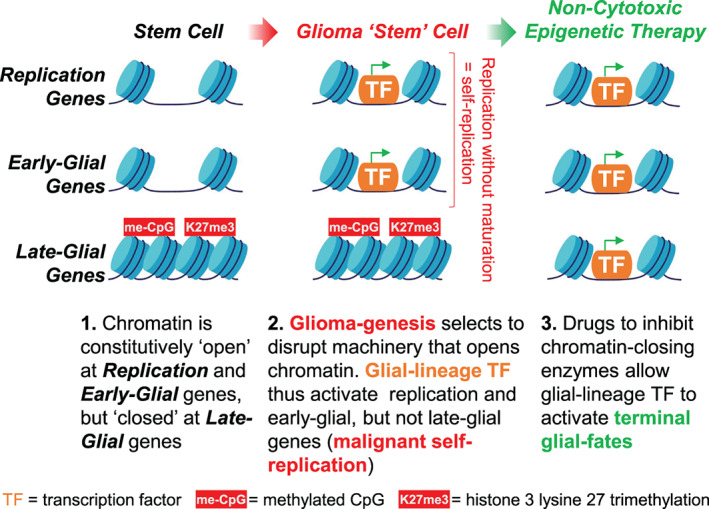
Glial-lineage malignancies (gliomas) recurrently mutate and/or delete the master regulators of apoptosis p53 and/or p16/CDKN2A, undermining apoptosis-intending (cytotoxic) treatments. By contrast to disrupted p53/p16, glioma cells are live-wired with the master transcription factor circuits that specify and drive glial lineage fates: these transcription factors activate early-glial and replication programs as expected, but fail in their other usual function of forcing onward glial lineage-maturation-late-glial genes have constitutively "closed" chromatin requiring chromatin-remodeling for activation-glioma-genesis disrupts several epigenetic components needed to perform this work, and simultaneously amplifies repressing epigenetic machinery instead. Pharmacologic inhibition of repressing epigenetic enzymes thus allows activation of late-glial genes and terminates glioma self-replication (self-replication = replication without lineage-maturation), independent of p53/p16/apoptosis. Lineage-specifying master transcription factors therefore contrast with p53/p16 in being enriched in self-replicating glioma cells, reveal a cause-effect relationship between aberrant epigenetic repression of late-lineage programs and malignant self-replication, and point to specific epigenetic targets for noncytotoxic glioma-therapy.
Keywords: cancer epigenetics; epigenetic; glioma; glioma therapy; neurooncology.
© 2022 The Authors. International Journal of Cancer published by John Wiley & Sons Ltd on behalf of UICC.
Conflict of interest statement
Ownership: Yogen Saunthararajah—EpiDestiny. Income: None. Research support: none. Intellectual property: Yogen Saunthararajah—patents around tetrahydrouridine, decitabine and 5‐azacytidine (US 9259469 B2; US 9265785 B2; US 9895391 B2) and cancer differentiation therapy (US 9926316 B2). Stefan Pusch—eligible to royalties as co‐inventor of BAY1436032. The corresponding patents are under the administrative supervision of the DKFZ technology transfer office. All other authors declare no conflicts of interest.
Figures
Similar articles
-
The early growth response gene EGR-1 behaves as a suppressor gene that is down-regulated independent of ARF/Mdm2 but not p53 alterations in fresh human gliomas.Clin Cancer Res. 2001 Sep;7(9):2788-96. Clin Cancer Res. 2001. PMID: 11555594
-
Introduction of wild-type p53 enhances thrombospondin-1 expression in human glioma cells.Cancer Lett. 2003 Feb 28;191(1):109-19. doi: 10.1016/s0304-3835(02)00592-x. Cancer Lett. 2003. PMID: 12609716
-
P53 gene mutation and expression of MDM2, P53, P16 protein and their relationship in human glioma.J Huazhong Univ Sci Technolog Med Sci. 2005;25(6):622-4, 635. doi: 10.1007/BF02896152. J Huazhong Univ Sci Technolog Med Sci. 2005. PMID: 16696307
-
[p16 tumor suppressor gene alteration in brain tumors].No To Shinkei. 1998 Jun;50(6):511-7. No To Shinkei. 1998. PMID: 9656246 Review. Japanese. No abstract available.
-
Gene therapy for gliomas: p53 and E2F-1 proteins and the target of apoptosis.Int J Mol Med. 1999 Jan;3(1):81-5. doi: 10.3892/ijmm.3.1.81. Int J Mol Med. 1999. PMID: 9864390 Review.
Cited by
-
Neuroendocrine lineage commitment of small cell lung cancers can be leveraged into p53-independent non-cytotoxic therapy.Cell Rep. 2023 Aug 29;42(8):113016. doi: 10.1016/j.celrep.2023.113016. Epub 2023 Aug 18. Cell Rep. 2023. PMID: 37597186 Free PMC article.
-
GQIcombi application to subdue glioma via differentiation therapy.Front Oncol. 2024 Jun 26;14:1322795. doi: 10.3389/fonc.2024.1322795. eCollection 2024. Front Oncol. 2024. PMID: 38988707 Free PMC article.
-
The Comparative Experimental Study of Sodium and Magnesium Dichloroacetate Effects on Pediatric PBT24 and SF8628 Cell Glioblastoma Tumors Using a Chicken Embryo Chorioallantoic Membrane Model and on Cells In Vitro.Int J Mol Sci. 2022 Sep 9;23(18):10455. doi: 10.3390/ijms231810455. Int J Mol Sci. 2022. PMID: 36142368 Free PMC article.
-
Oncotherapy resistance explained by Darwinian and Lamarckian models.J Clin Invest. 2024 Apr 15;134(8):e179788. doi: 10.1172/JCI179788. J Clin Invest. 2024. PMID: 38618954 Free PMC article.
References
-
- Mukherjee S. The Emperor of all Maladies: a Biography of Cancer. 1st Scribner hardcover ed. New York: Scribner; 2010. xiv, 571 p., 8 p. of plates.
-
- Velcheti V, Schrump D, Saunthararajah Y. Ultimate precision: targeting cancer but not Normal self‐replication. Am Soc Clin Oncol Educ Book. 2018;38:950‐963. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
