

Consecutive action of two BAHD acyltransferases promotes tetracoumaroyl spermine accumulation in chicory
- PMID: 35604091
- PMCID: PMC9343010
- DOI: 10.1093/plphys/kiac234
Consecutive action of two BAHD acyltransferases promotes tetracoumaroyl spermine accumulation in chicory
Abstract
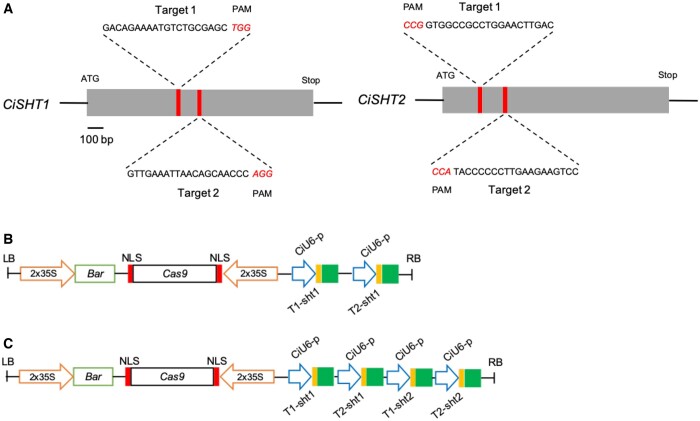
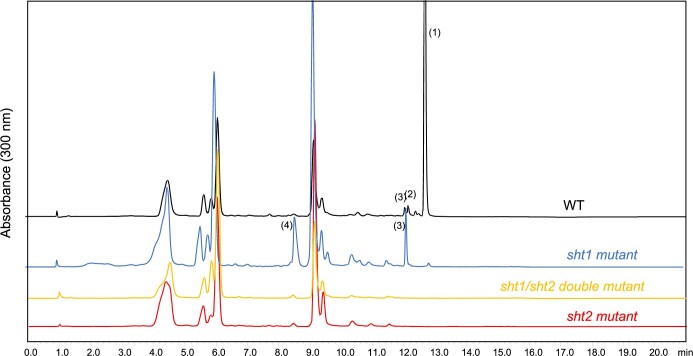
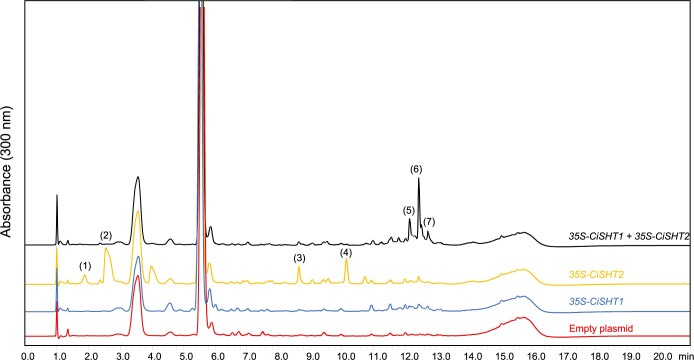
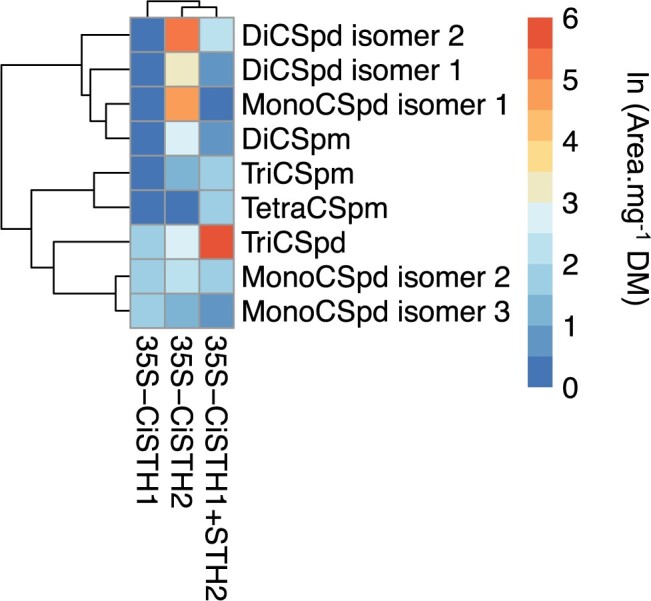
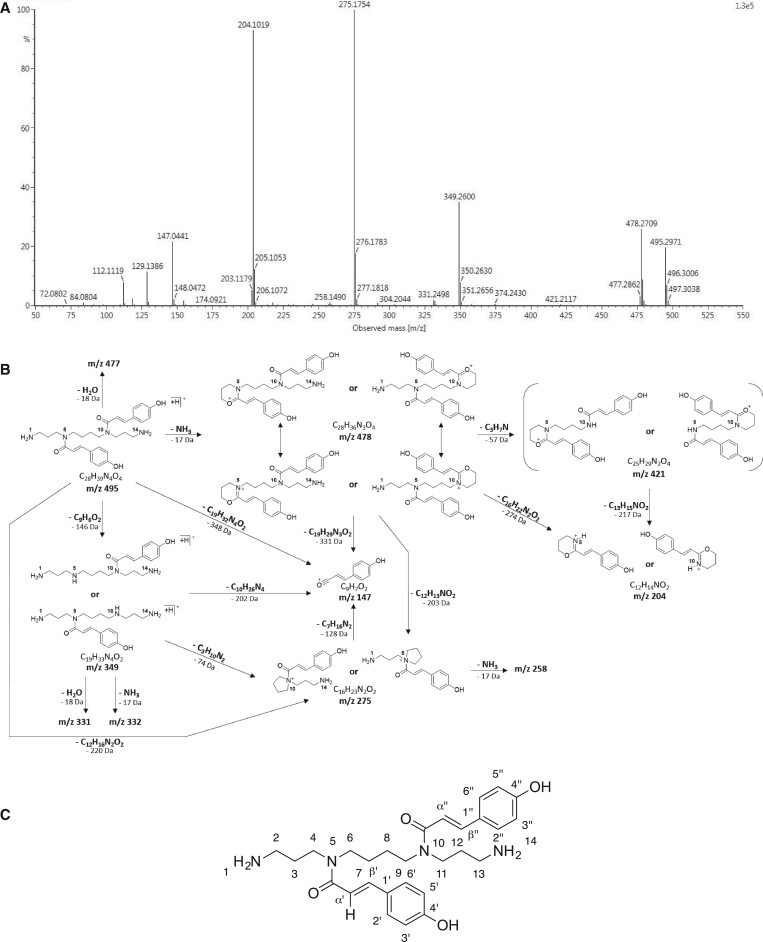
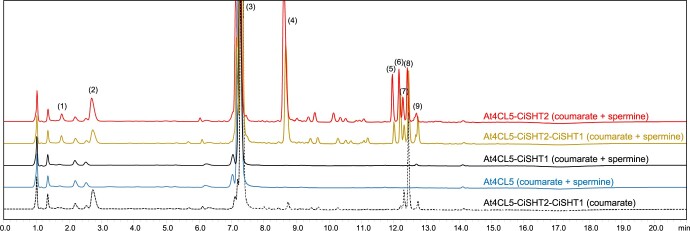
Fully substituted phenolamide accumulation in the pollen coat of Eudicotyledons is a conserved evolutionary chemical trait. Interestingly, spermidine derivatives are replaced by spermine derivatives as the main phenolamide accumulated in the Asteraceae family. Here, we show that the full substitution of spermine in chicory (Cichorium intybus) requires the successive action of two enzymes, that is spermidine hydroxycinnamoyl transferase-like proteins 1 and 2 (CiSHT1 and CiSHT2), two members of the BAHD enzyme family. Deletion of these genes in chicory using CRISPR/Cas9 gene editing technology evidenced that CiSHT2 catalyzes the first N-acylation steps, whereas CiSHT1 fulfills the substitution to give rise to tetracoumaroyl spermine. Additional experiments using Nicotiana benthamiana confirmed these findings. Expression of CiSHT2 alone promoted partially substituted spermine accumulation, and coexpression of CiSHT2 and CiSHT1 promoted synthesis and accumulation of the fully substituted spermine. Structural characterization of the main product of CiSHT2 using nuclear magnetic resonance revealed that CiSHT2 preferentially catalyzed N-acylation of secondary amines to form N5,N10-dicoumaroyl spermine, whereas CiSHT1 used this substrate to synthesize tetracoumaroyl spermine. We showed that spermine availability may be a key determinant toward preferential accumulation of spermine derivatives over spermidine derivatives in chicory. Our results reveal a subfunctionalization among the spermidine hydroxycinnamoyl transferase that was accompanied by a modification of free polyamine metabolism that has resulted in the accumulation of this new phenolamide in chicory and most probably in all Asteraceae. Finally, genetically engineered yeast (Saccharomyces cerevisiae) was shown to be a promising host platform to produce these compounds.
© The Author(s) 2022. Published by Oxford University Press on behalf of American Society of Plant Biologists.
Figures
References
-
- Ahmed E, Arshad M, Zakriyya Khan M, Shoaib Amjad M, Mehreen Sadaf H, Riaz I, Sidra Sabir P, Ahmad N, Sabaoon P, Ejaz Ahmed P, et al. (2017) Secondary metabolites and their multidimensional prospective in plant life. J Pharmacogn Phytochem 6: 205–214
-
- Bassard JE, Ullmann P, Bernier F, Werck-Reichhart D (2010) Phenolamides: bridging polyamines to the phenolic metabolism. Phytochemistry 71: 1808–1824 - PubMed
-
- Bigler L, Schnider CF, Hu W, Hesse M (1996) Acid-catalyzed isomerization of N,N’-Bis[(E)-3-(4-hydroxyphenyl)prop-2-enoyl]spermidines by the zip reaction. Helv Chim Acta 79: 2152–2163
-
- Bogdanovic MD, Todorovic SI, Banjanac T, Dragiƒáeviƒá MB, Verstappen FWA, Bouwmeester HJ, Simonoviƒá AD, Bogdanović MD, Todorović SI, Banjanac T, et al. (2014) Production of guaianolides in Agrobacterium rhizogenes - transformed chicory regenerants flowering in vitro. Ind Crops Prod 60: 52–59
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
