

Calpain-2 Mediates MBNL2 Degradation and a Developmental RNA Processing Program in Neurodegeneration
- PMID: 35606145
- PMCID: PMC9233439
- DOI: 10.1523/JNEUROSCI.2006-21.2022
Calpain-2 Mediates MBNL2 Degradation and a Developmental RNA Processing Program in Neurodegeneration
Abstract
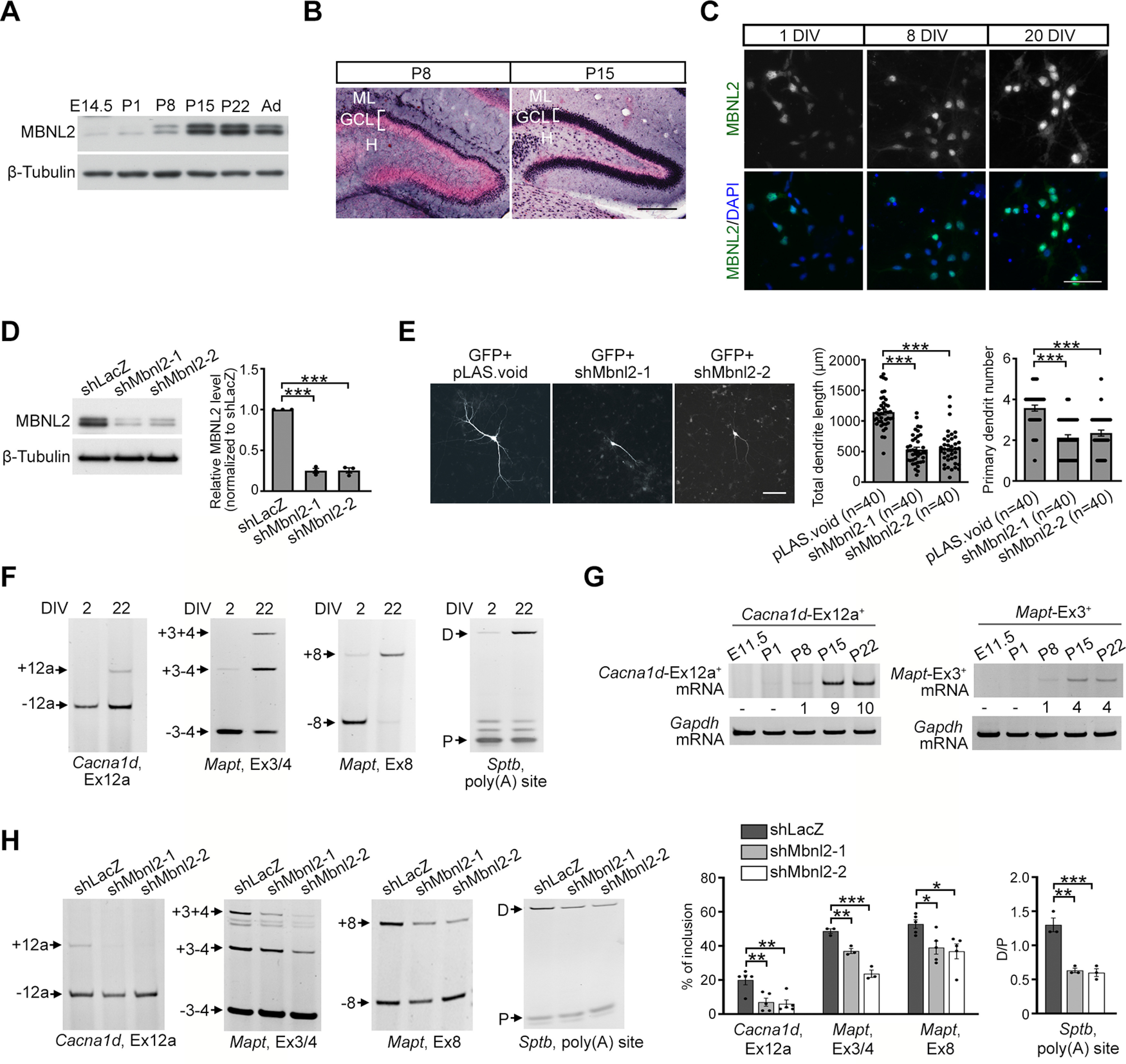
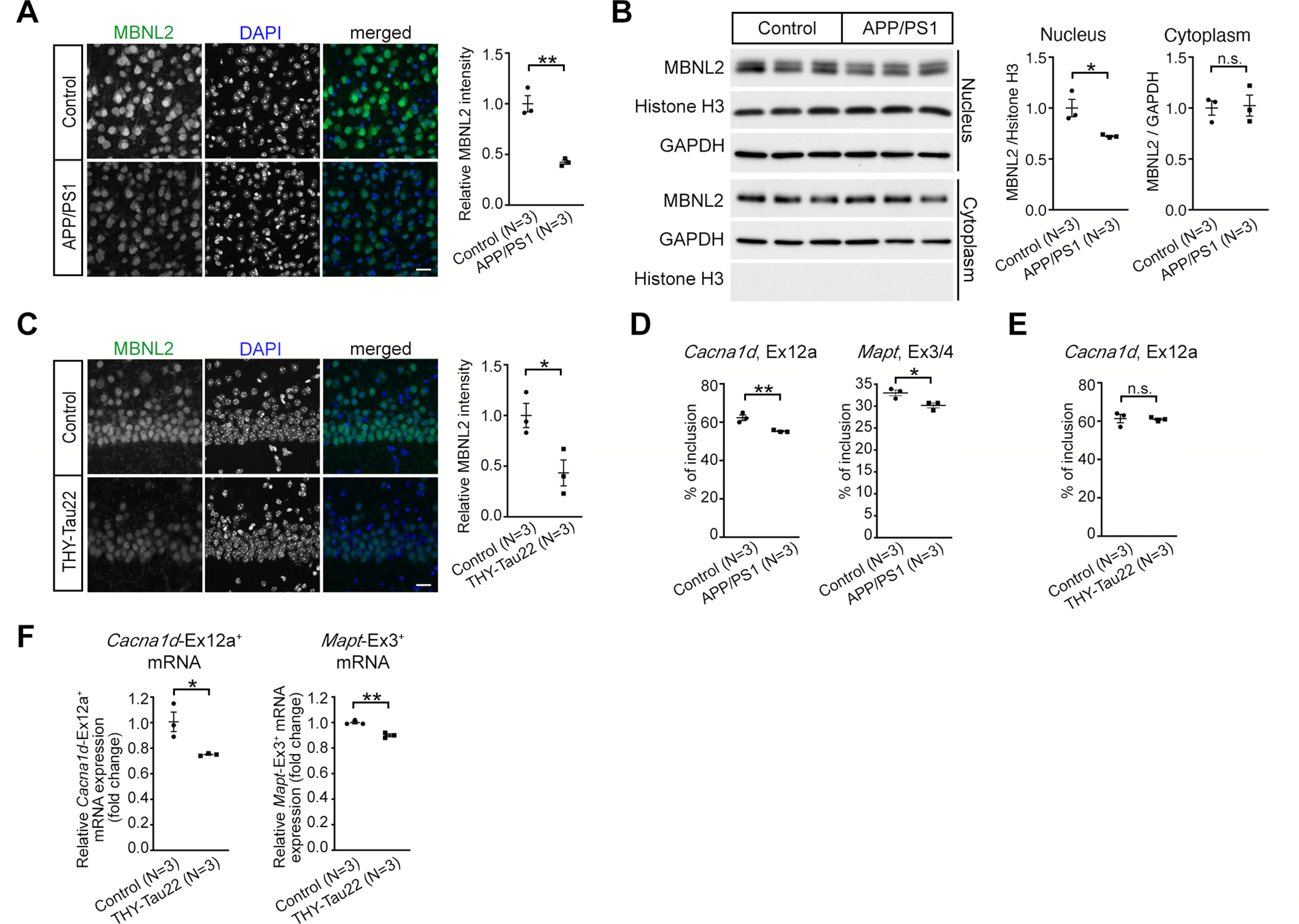
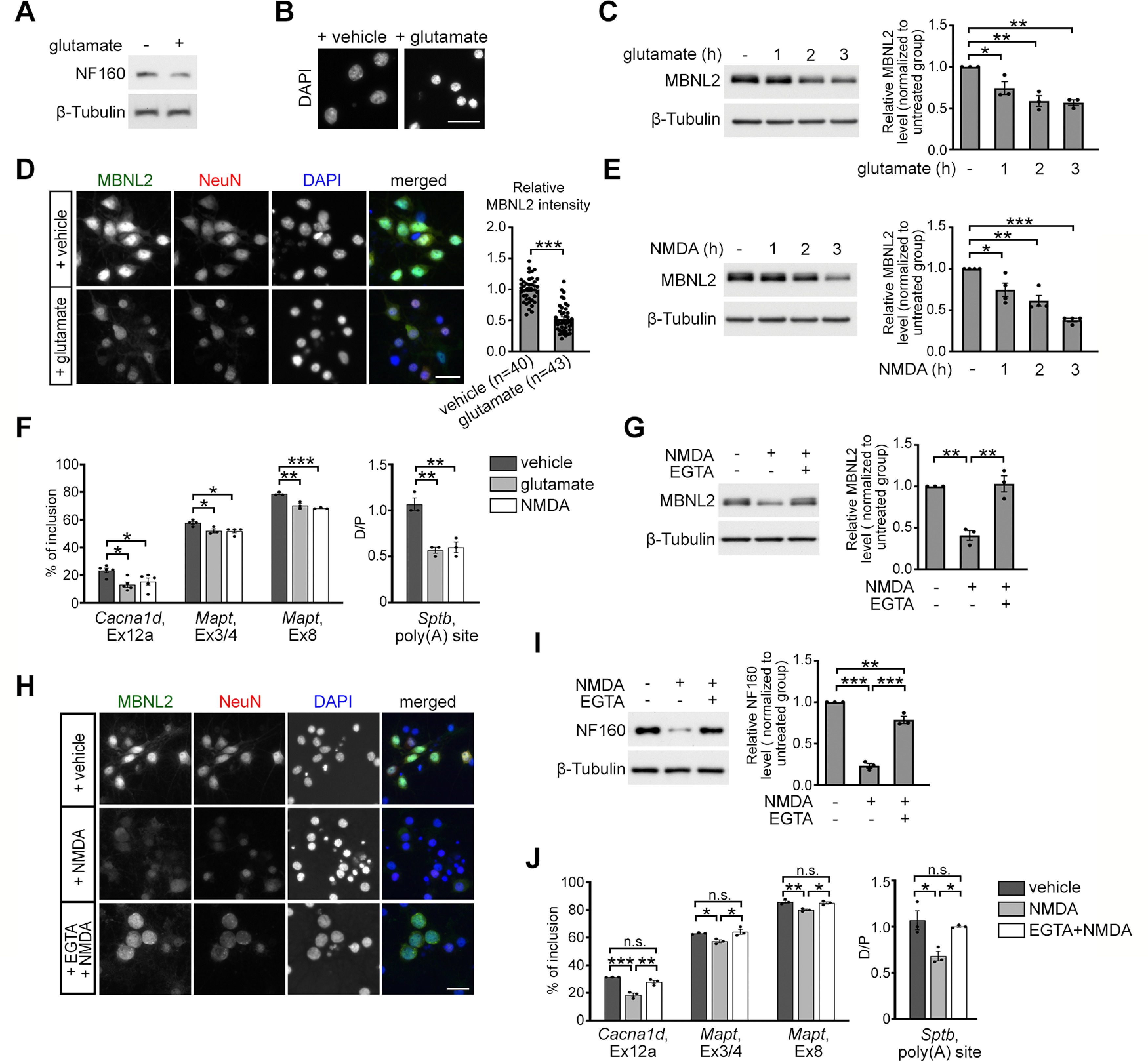
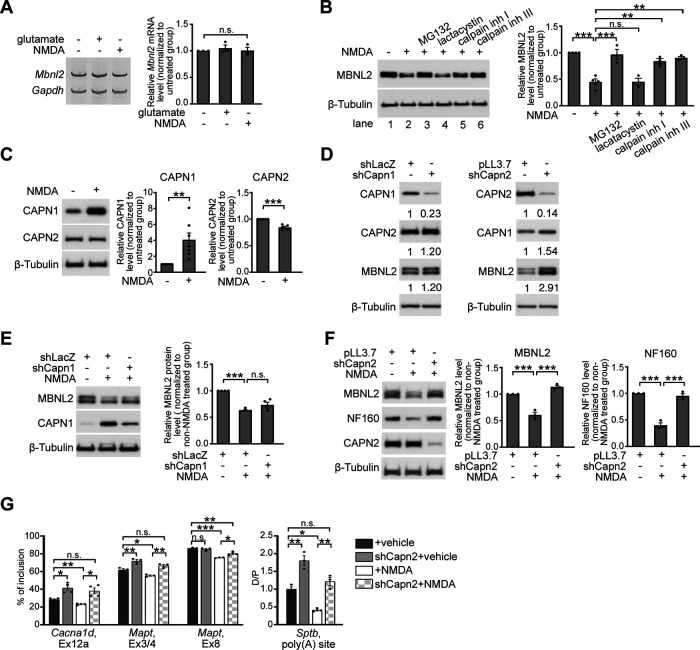
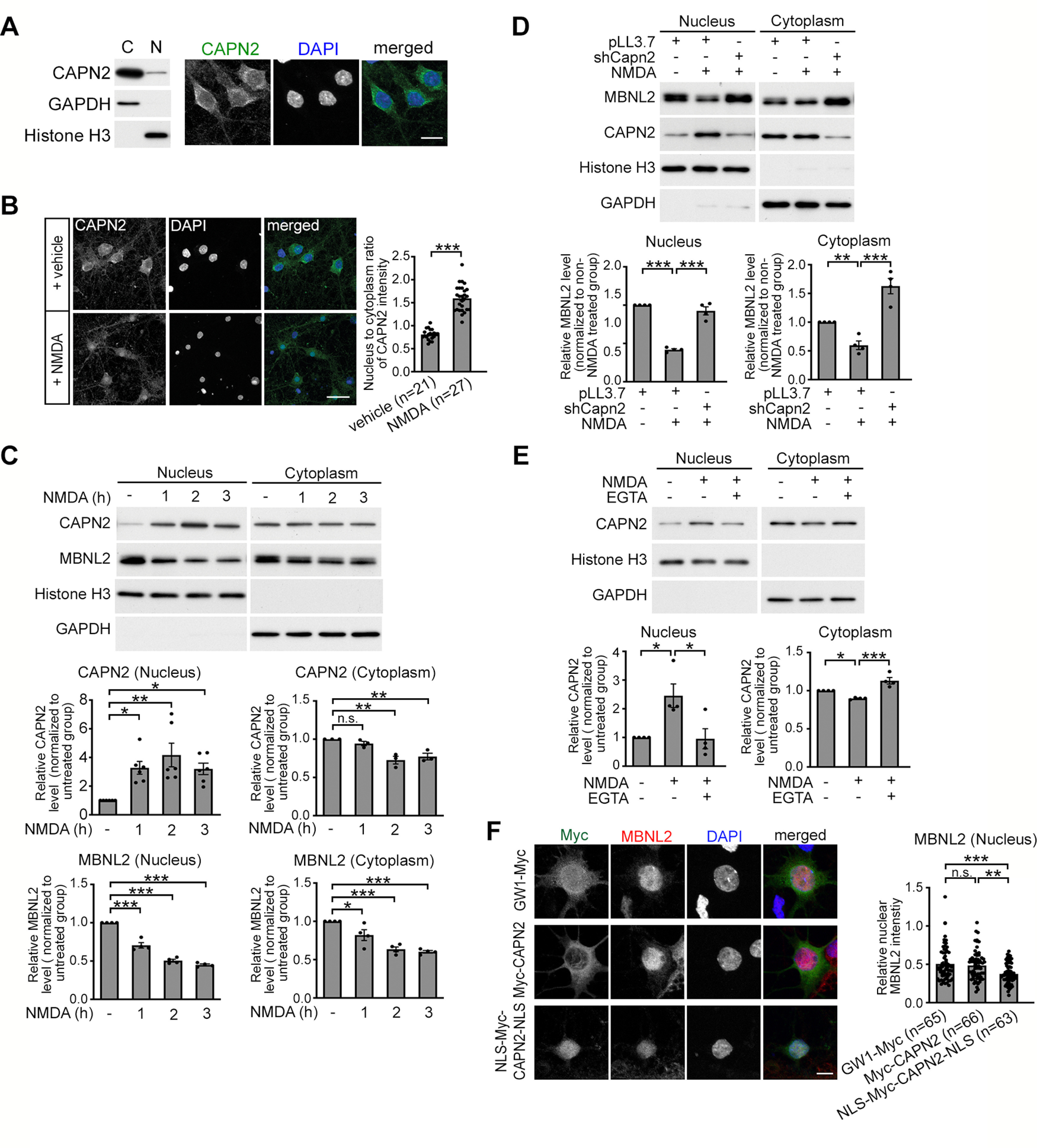
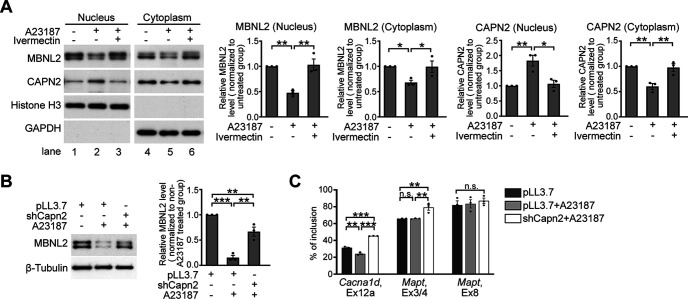
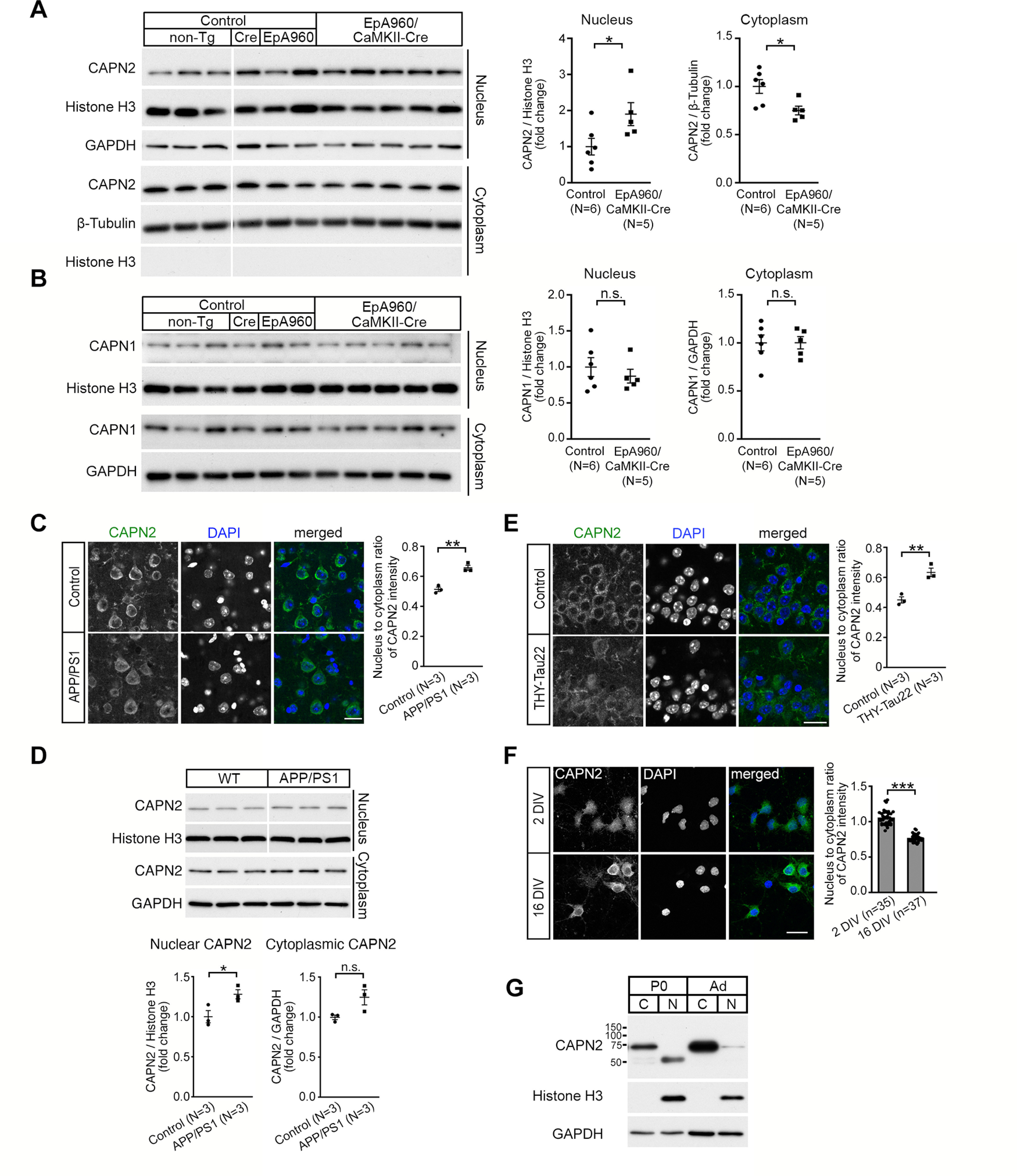
Increasing loss of structure and function of neurons and decline in cognitive function is commonly seen during the progression of neurologic diseases, although the causes and initial symptoms of individual diseases are distinct. This observation suggests a convergence of common degenerative features. In myotonic dystrophy type 1 (DM1), the expression of expanded CUG RNA induces neurotransmission dysfunction before axon and dendrite degeneration and reduced MBNL2 expression associated with aberrant alternative splicing. The role of loss of function of MBNL2 in the pathogenesis of neurodegeneration and the causal mechanism of neurodegeneration-reduced expression of MBNL2 remain elusive. Here, we show that increased MBNL2 expression is associated with neuronal maturation and required for neuronal morphogenesis and the fetal to adult developmental transition of RNA processing. Neurodegenerative conditions including NMDA receptor (NMDAR)-mediated excitotoxicity and dysregulated calcium homeostasis triggered nuclear translocation of calpain-2, thus resulting in MBNL2 degradation and reversal of MBNL2-regulated RNA processing to developmental patterns. Nuclear expression of calpain-2 resembled its developmental pattern and was associated with MBNL2 degradation. Knock-down of calpain-2 expression or inhibition of calpain-2 nuclear translocation prevented neurodegeneration-reduced MBNL2 expression and dysregulated RNA processing. Increased calpain-2 nuclear translocation associated with reduced MBNL2 expression and aberrant RNA processing occurred in models for DM1 and Alzheimer's disease (AD) including EpA960/CaMKII-Cre mice of either sex and female APP/PS1 and THY-Tau22 mice. Our results identify a regulatory mechanism for MBNL2 downregulation and suggest that calpain-2-mediated MBNL2 degradation accompanied by re-induction of a developmental RNA processing program may be a converging pathway to neurodegeneration.SIGNIFICANCE STATEMENT Neurologic diseases share many features during disease progression, such as cognitive decline and brain atrophy, which suggests a common pathway for developing degenerative features. Here, we show that the neurodegenerative conditions glutamate-induced excitotoxicity and dysregulated calcium homeostasis induced translocation of the cysteine protease calpain-2 into the nucleus, resulting in MBNL2 degradation and reversal of MBNL2-regulated RNA processing to an embryonic pattern. Knock-down or inhibition of nuclear translocation of calpain-2 prevented MBNL2 degradation and maintained MBNL2-regulated RNA processing in the adult pattern. Models of myotonic dystrophy and Alzheimer's disease (AD) also showed calpain-2-mediated MBNL2 degradation and a developmental RNA processing program. Our studies suggest MBNL2 function disrupted by calpain-2 as a common pathway, thus providing an alternative therapeutic strategy for neurodegeneration.
Keywords: Alzheimer's disease; MBNL2; RNA-processing; calpain-2; excitotoxicity; myotonic dystrophy.
Copyright © 2022 the authors.
Figures
Similar articles
-
MBNL1 is the primary determinant of focus formation and aberrant insulin receptor splicing in DM1.J Biol Chem. 2005 Feb 18;280(7):5773-80. doi: 10.1074/jbc.M410781200. Epub 2004 Nov 16. J Biol Chem. 2005. PMID: 15546872
-
Choroid plexus mis-splicing and altered cerebrospinal fluid composition in myotonic dystrophy type 1.Brain. 2023 Oct 3;146(10):4217-4232. doi: 10.1093/brain/awad148. Brain. 2023. PMID: 37143315 Free PMC article.
-
Reduced cytoplasmic MBNL1 is an early event in a brain-specific mouse model of myotonic dystrophy.Hum Mol Genet. 2017 Jun 15;26(12):2247-2257. doi: 10.1093/hmg/ddx115. Hum Mol Genet. 2017. PMID: 28369378
-
An Overview of Alternative Splicing Defects Implicated in Myotonic Dystrophy Type I.Genes (Basel). 2020 Sep 22;11(9):1109. doi: 10.3390/genes11091109. Genes (Basel). 2020. PMID: 32971903 Free PMC article. Review.
-
Pathogenic mechanisms of myotonic dystrophy.Biochem Soc Trans. 2009 Dec;37(Pt 6):1281-6. doi: 10.1042/BST0371281. Biochem Soc Trans. 2009. PMID: 19909263 Free PMC article. Review.
Cited by
-
Functions of the Muscleblind-like protein family and their role in disease.Cell Commun Signal. 2025 Feb 18;23(1):97. doi: 10.1186/s12964-025-02102-5. Cell Commun Signal. 2025. PMID: 39966885 Free PMC article. Review.
-
Loss of MBNL1-mediated retrograde BDNF signaling in the myotonic dystrophy brain.Acta Neuropathol Commun. 2023 Mar 15;11(1):44. doi: 10.1186/s40478-023-01540-x. Acta Neuropathol Commun. 2023. PMID: 36922901 Free PMC article.
-
Cognitive impairment, neuroimaging abnormalities, and their correlations in myotonic dystrophy: a comprehensive review.Front Cell Neurosci. 2024 Apr 4;18:1369332. doi: 10.3389/fncel.2024.1369332. eCollection 2024. Front Cell Neurosci. 2024. PMID: 38638300 Free PMC article. Review.
-
Construction of a Novel Necroptosis-Related Signature in Rat DRG for Neuropathic Pain.J Inflamm Res. 2025 Jan 6;18:147-165. doi: 10.2147/JIR.S494286. eCollection 2025. J Inflamm Res. 2025. PMID: 39802520 Free PMC article.
-
CAPN2 correlates with insulin resistance states in PCOS as evidenced by multi-dataset analysis.J Ovarian Res. 2024 Apr 12;17(1):79. doi: 10.1186/s13048-024-01407-2. J Ovarian Res. 2024. PMID: 38610028 Free PMC article.
References
-
- Batra R, Charizanis K, Manchanda M, Mohan A, Li M, Finn DJ, Goodwin M, Zhang C, Sobczak K, Thornton CA, Swanson MS (2014) Loss of MBNL leads to disruption of developmentally regulated alternative polyadenylation in RNA-mediated disease. Mol Cell 56:311–322. 10.1016/j.molcel.2014.08.027 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases