

Genome of the parasitoid wasp Cotesia chilonis sheds light on amino acid resource exploitation
- PMID: 35606775
- PMCID: PMC9128236
- DOI: 10.1186/s12915-022-01313-3
Genome of the parasitoid wasp Cotesia chilonis sheds light on amino acid resource exploitation
Abstract
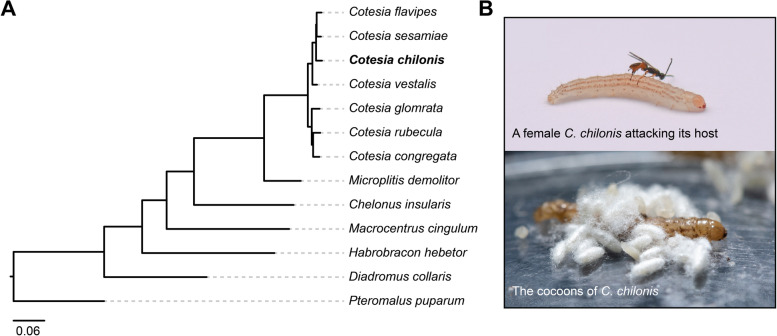
Background: A fundamental feature of parasitism is the nutritional exploitation of host organisms by their parasites. Parasitoid wasps lay eggs on arthropod hosts, exploiting them for nutrition to support larval development by using diverse effectors aimed at regulating host metabolism. However, the genetic components and molecular mechanisms at the basis of such exploitation, especially the utilization of host amino acid resources, remain largely unknown. To address this question, here, we present a chromosome-level genome assembly of the parasitoid wasp Cotesia chilonis and reconstruct its amino acid biosynthetic pathway.
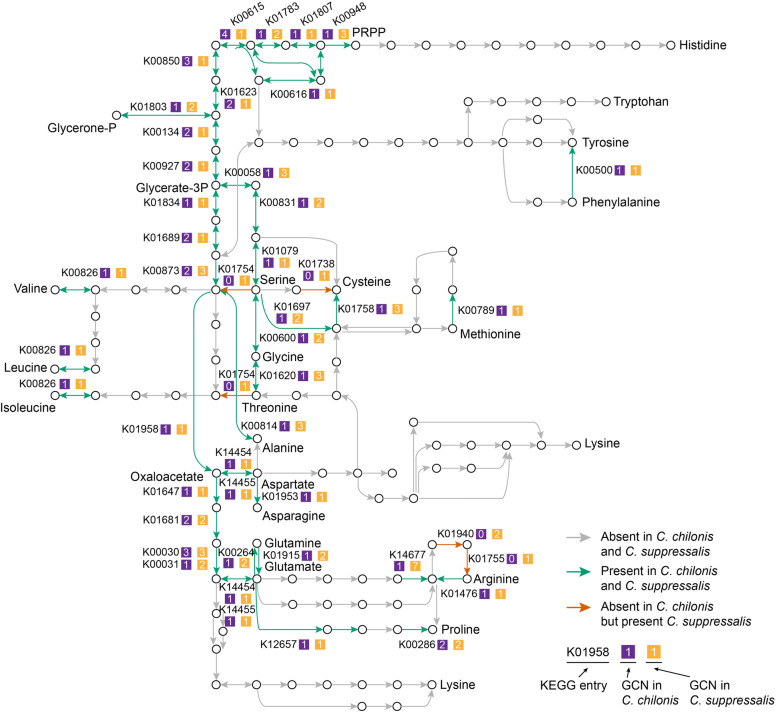
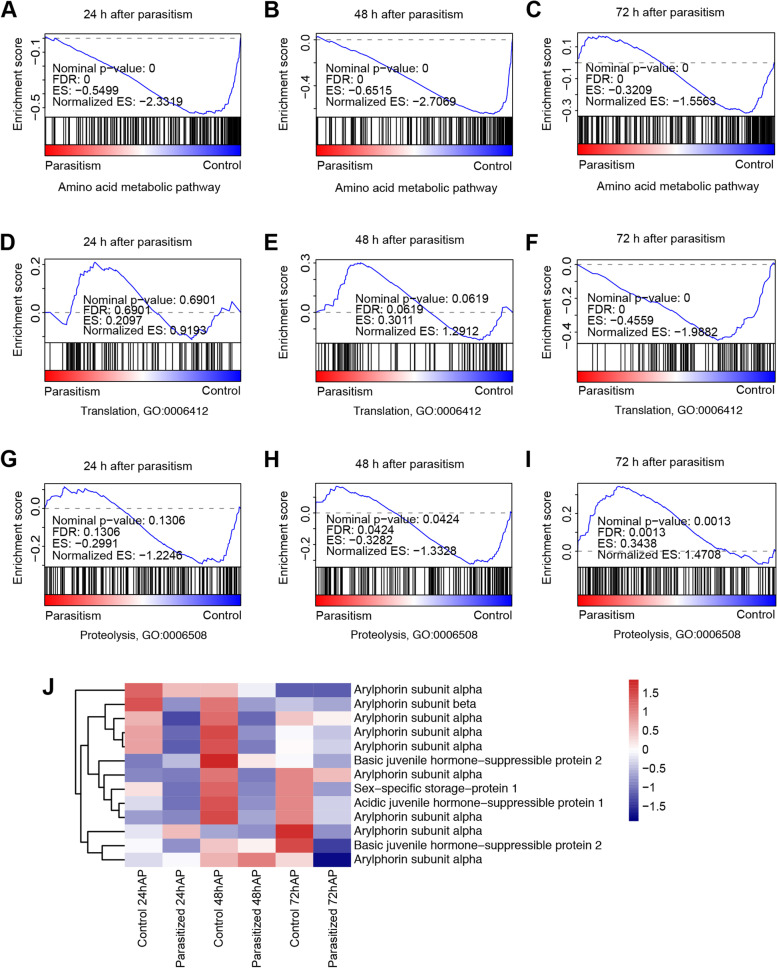
Results: Analyses of the amino acid synthetic pathway indicate that C. chilonis lost the ability to synthesize ten amino acids, which was confirmed by feeding experiments with amino acid-depleted media. Of the ten pathways, nine are known to have been lost in the common ancestor of animals. We find that the ability to synthesize arginine was also lost in C. chilonis because of the absence of two key genes in the arginine synthesis pathway. Further analyses of the genomes of 72 arthropods species show that the loss of arginine synthesis is common in arthropods. Metabolomic analyses by UPLC-MS/MS reveal that the temporal concentrations of arginine, serine, tyrosine, and alanine are significantly higher in host (Chilo suppressalis) hemolymph at 3 days after parasitism, whereas the temporal levels of 5-hydroxylysine, glutamic acid, methionine, and lysine are significantly lower. We sequence the transcriptomes of a parasitized host and non-parasitized control. Differential gene expression analyses using these transcriptomes indicate that parasitoid wasps inhibit amino acid utilization and activate protein degradation in the host, likely resulting in the increase of amino acid content in host hemolymph.
Conclusions: We sequenced the genome of a parasitoid wasp, C. chilonis, and revealed the features of trait loss in amino acid biosynthesis. Our work provides new insights into amino acid exploitation by parasitoid wasps, and this knowledge can specifically be used to design parasitoid artificial diets that potentially benefit mass rearing of parasitoids for pest control.
Keywords: Amino acid synthesis; Cotesia chilonis; Genome sequencing; Nutrition exploitation; Parasitoid wasps; Trait loss.
© 2022. The Author(s).
Conflict of interest statement
The authors declare that they have no competing interests.
Figures



Similar articles
-
Amino acid synthesis loss in parasitoid wasps and other hymenopterans.Elife. 2020 Oct 19;9:e59795. doi: 10.7554/eLife.59795. Elife. 2020. Retraction in: Elife. 2020 Dec 01;9:e65123. doi: 10.7554/eLife.65123. PMID: 33074103 Free PMC article. Retracted.
-
Screening Sugars Can Benefit the Parasitoid Cotesia chilonis (Hymenoptera: Braconidae) Without Benefiting Its Host, Chilo suppressalis (Lepidoptera: Crambidae).J Econ Entomol. 2019 Sep 23;112(5):2142-2148. doi: 10.1093/jee/toz166. J Econ Entomol. 2019. PMID: 31219581
-
Transcriptome analysis of an endoparasitoid wasp Cotesia chilonis (Hymenoptera: Braconidae) reveals genes involved in successful parasitism.Arch Insect Biochem Physiol. 2015 Apr;88(4):203-21. doi: 10.1002/arch.21214. Epub 2014 Oct 22. Arch Insect Biochem Physiol. 2015. PMID: 25336406
-
Role of virus-like particles in parasitoid-host interaction of insects.Subcell Biochem. 1989;15:91-119. doi: 10.1007/978-1-4899-1675-4_4. Subcell Biochem. 1989. PMID: 2678620 Review.
-
The state of parasitoid wasp genomics.Trends Parasitol. 2024 Oct;40(10):914-929. doi: 10.1016/j.pt.2024.08.003. Epub 2024 Sep 2. Trends Parasitol. 2024. PMID: 39227194 Review.
Cited by
-
Stage-specific metabolic allocation: nutrient investment strategies during Lysiphlebia japonica Ashmead development.Front Nutr. 2025 Aug 6;12:1636519. doi: 10.3389/fnut.2025.1636519. eCollection 2025. Front Nutr. 2025. PMID: 40843203 Free PMC article.
-
Host and venom evolution in parasitoid wasps: does independently adapting to the same host shape the evolution of the venom gland transcriptome?BMC Biol. 2024 Aug 15;22(1):174. doi: 10.1186/s12915-024-01974-2. BMC Biol. 2024. PMID: 39148049 Free PMC article.
-
Chromosome-level genome assembly of the bethylid ectoparasitoid wasp Sclerodermus sp. 'alternatusi'.Sci Data. 2024 May 2;11(1):438. doi: 10.1038/s41597-024-03278-0. Sci Data. 2024. PMID: 38698068 Free PMC article.
-
Integration of transcriptomic and proteomic data uncovers the nutritional requirement of the immature development of an endoparasitoid wasp.Sci Rep. 2025 May 22;15(1):17744. doi: 10.1038/s41598-025-02290-7. Sci Rep. 2025. PMID: 40404784 Free PMC article.
-
Evaluating Beauveria bassiana Strains for Insect Pest Control and Endophytic Colonization in Wheat.Insects. 2025 Mar 10;16(3):287. doi: 10.3390/insects16030287. Insects. 2025. PMID: 40266821 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
