

Glomerular basement membrane deposition of collagen α1(III) in Alport glomeruli by mesangial filopodia injures podocytes via aberrant signaling through DDR1 and integrin α2β1
- PMID: 35607980
- PMCID: PMC9378723
- DOI: 10.1002/path.5969
Glomerular basement membrane deposition of collagen α1(III) in Alport glomeruli by mesangial filopodia injures podocytes via aberrant signaling through DDR1 and integrin α2β1
Abstract
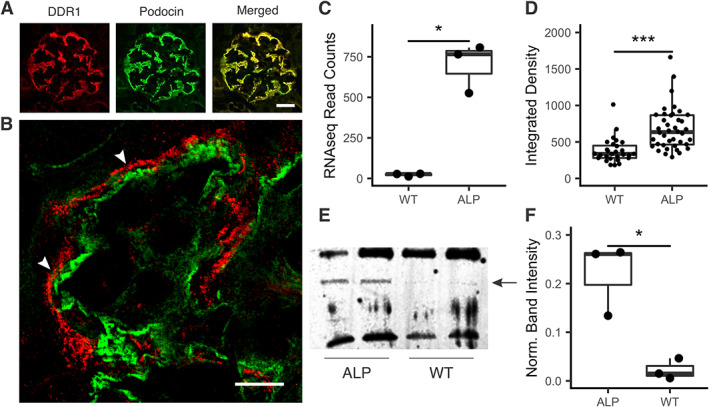
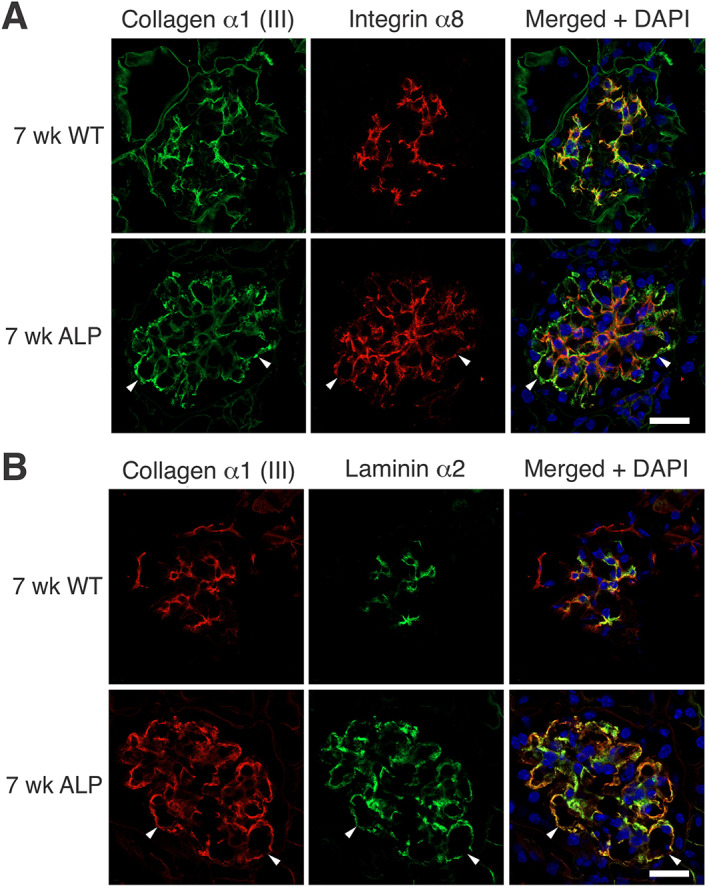
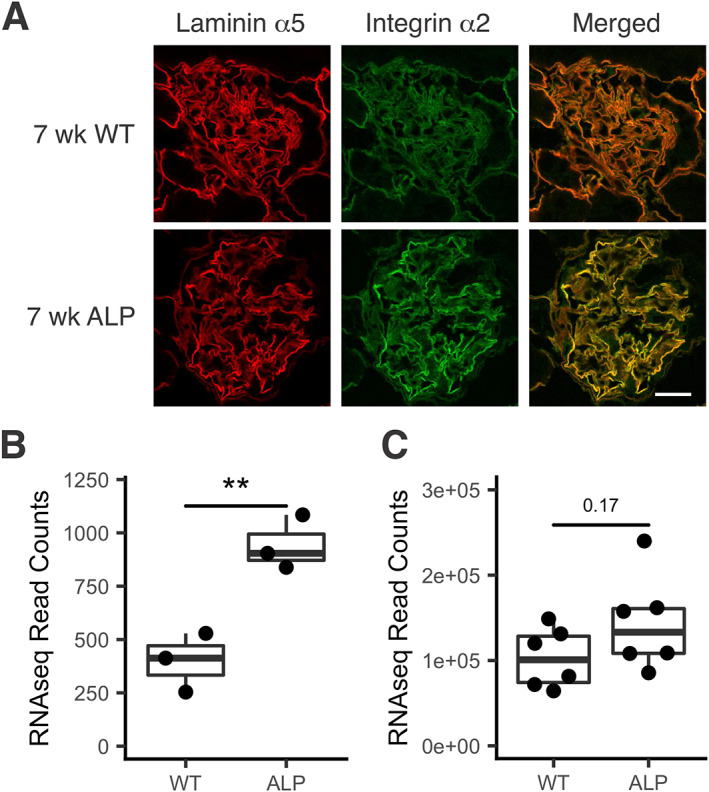
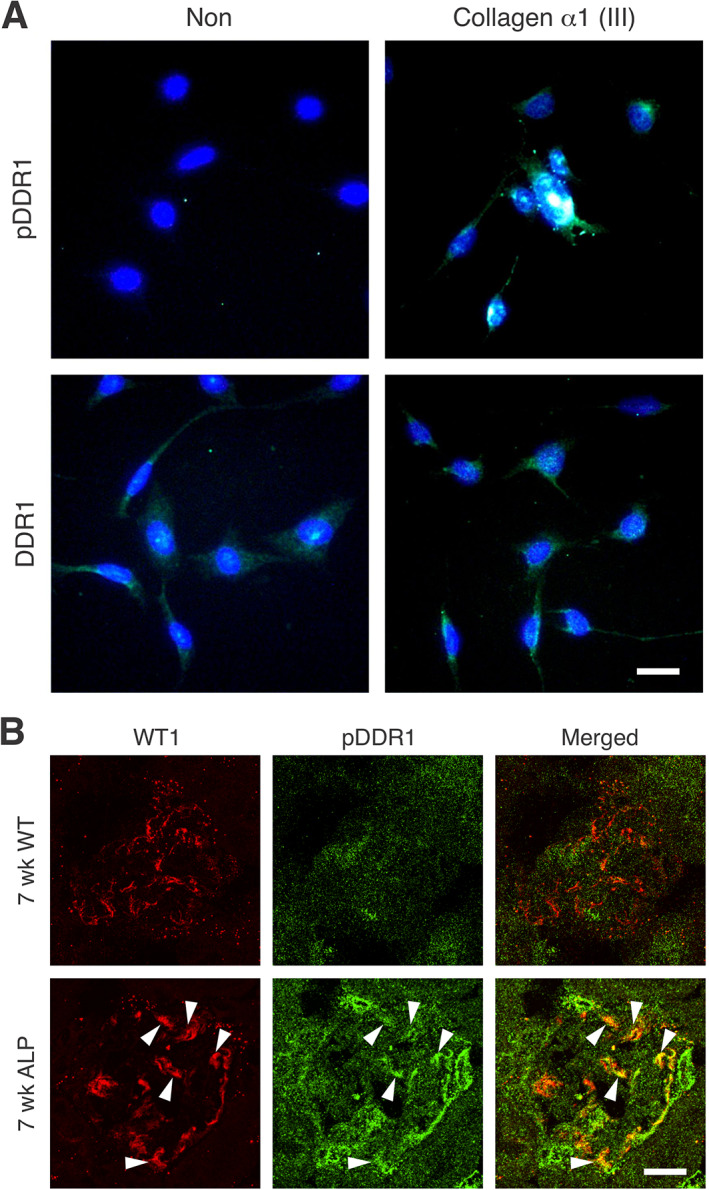
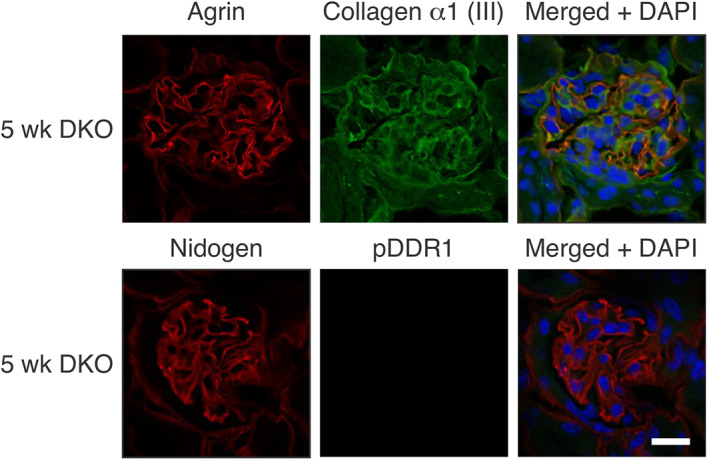
In Alport mice, activation of the endothelin A receptor (ETA R) in mesangial cells results in sub-endothelial invasion of glomerular capillaries by mesangial filopodia. Filopodia deposit mesangial matrix in the glomerular basement membrane (GBM), including laminin 211 which activates NF-κB, resulting in induction of inflammatory cytokines. Herein we show that collagen α1(III) is also deposited in the GBM. Collagen α1(III) localized to the mesangium in wild-type mice and was found in both the mesangium and the GBM in Alport mice. We show that collagen α1(III) activates discoidin domain receptor family, member 1 (DDR1) receptors both in vitro and in vivo. To elucidate whether collagen α1(III) might cause podocyte injury, cultured murine Alport podocytes were overlaid with recombinant collagen α1(III), or not, for 24 h and RNA was analyzed by RNA sequencing (RNA-seq). These same cells were subjected to siRNA knockdown for integrin α2 or DDR1 and the RNA was analyzed by RNA-seq. Results were validated in vivo using RNA-seq from RNA isolated from wild-type and Alport mouse glomeruli. Numerous genes associated with podocyte injury were up- or down-regulated in both Alport glomeruli and cultured podocytes treated with collagen α1(III), 18 of which have been associated previously with podocyte injury or glomerulonephritis. The data indicate α2β1 integrin/DDR1 co-receptor signaling as the dominant regulatory mechanism. This may explain earlier studies where deletion of either DDR1 or α2β1 integrin in Alport mice ameliorates renal pathology. © 2022 Boys Town National Research Hospital. The Journal of Pathology published by John Wiley & Sons Ltd on behalf of The Pathological Society of Great Britain and Ireland.
Keywords: Alport syndrome; collagen α1(III); discoidin domain receptor 1; integrin α2β1; podocyte injury.
© 2022 Boys Town National Research Hospital. The Journal of Pathology published by John Wiley & Sons Ltd on behalf of The Pathological Society of Great Britain and Ireland.
Figures
References
-
- Mochizuki T, Lemmink HH, Mariyama M, et al. Identification of mutations in the α3(IV) and α4(IV) collagen genes in autosomal recessive Alport syndrome. Nat Genet 1994; 8: 77–82. - PubMed
-
- Lemmink HH, Mochizuki T, van den Heuvel LP, et al. Mutations in the type IV collagen alpha 3 (COL4A3) gene in autosomal recessive Alport syndrome. Hum Mol Genet 1994; 3: 1269–1273. - PubMed
-
- Barker DF, Hostikka SL, Zhou J, et al. Identification of mutations in the COL4A5 collagen gene in Alport syndrome. Science 1990; 248: 1224–1227. - PubMed
-
- Gunwar S, Ballester F, Noelken ME, et al. Glomerular basement membrane. Identification of a novel disulfide‐cross‐linked network of α3, α4, and α5 chains of type IV collagen and its implications for the pathogenesis of Alport syndrome. J Biol Chem 1998; 273: 8767–8775. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
