

The neuronal tyrosine kinase receptor ligand ALKAL2 mediates persistent pain
- PMID: 35608912
- PMCID: PMC9197515
- DOI: 10.1172/JCI154317
The neuronal tyrosine kinase receptor ligand ALKAL2 mediates persistent pain
Abstract
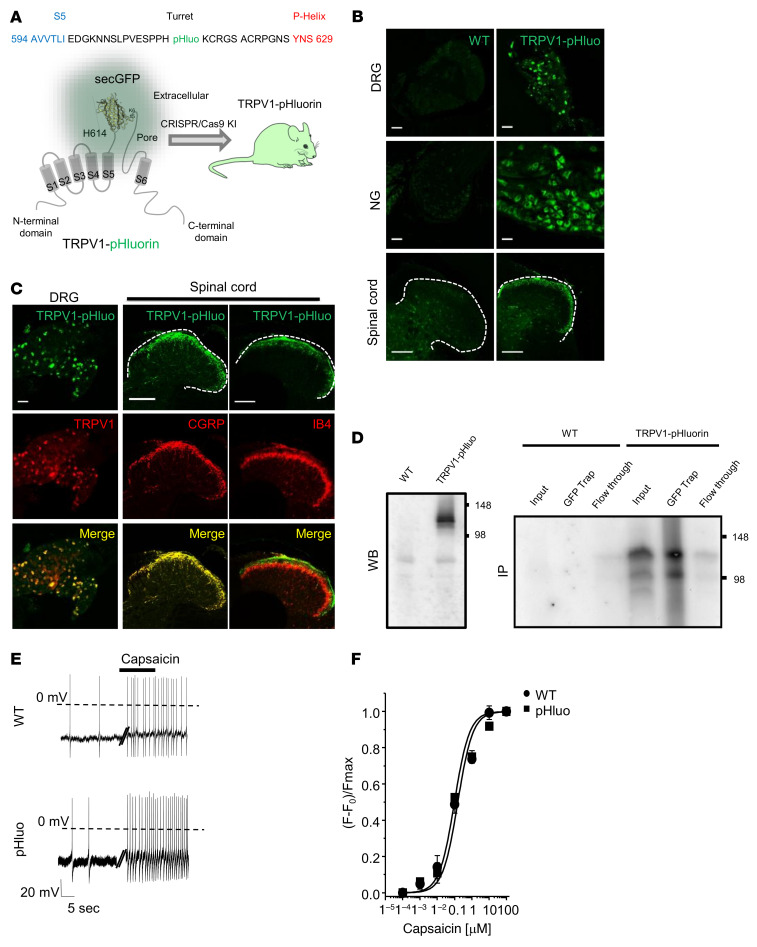
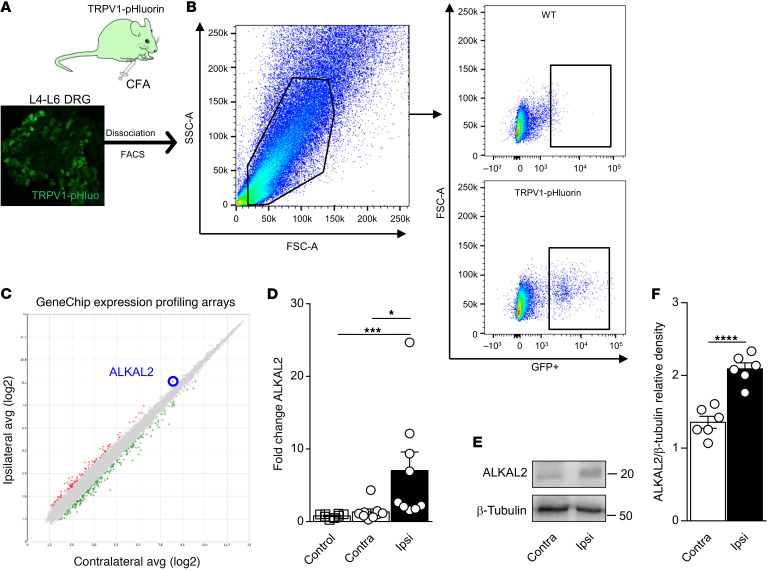
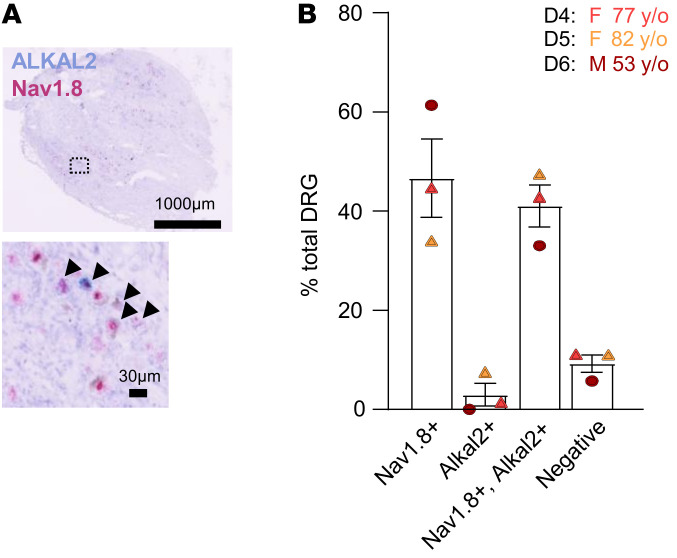
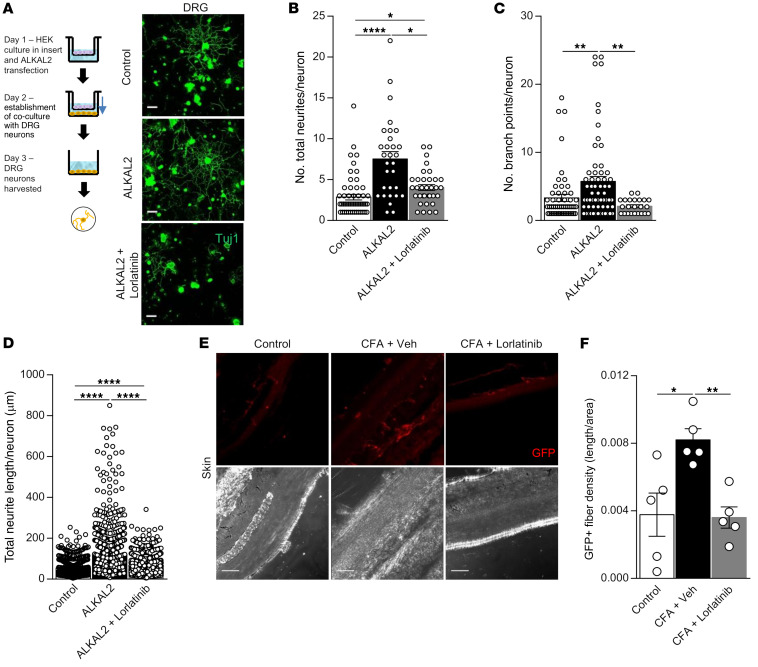
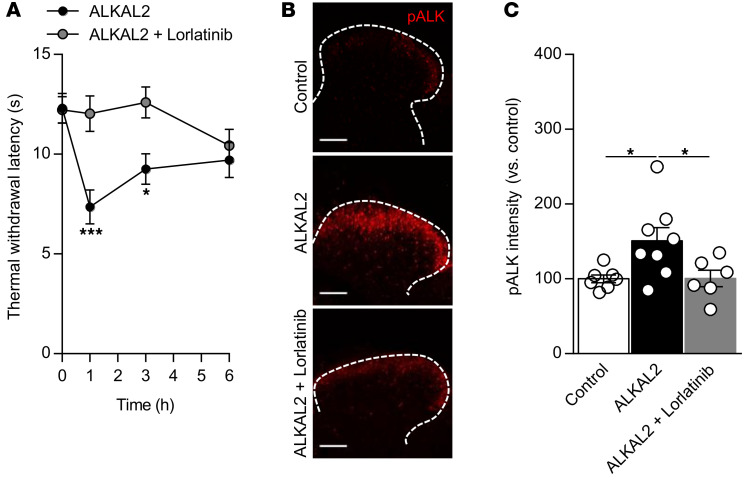
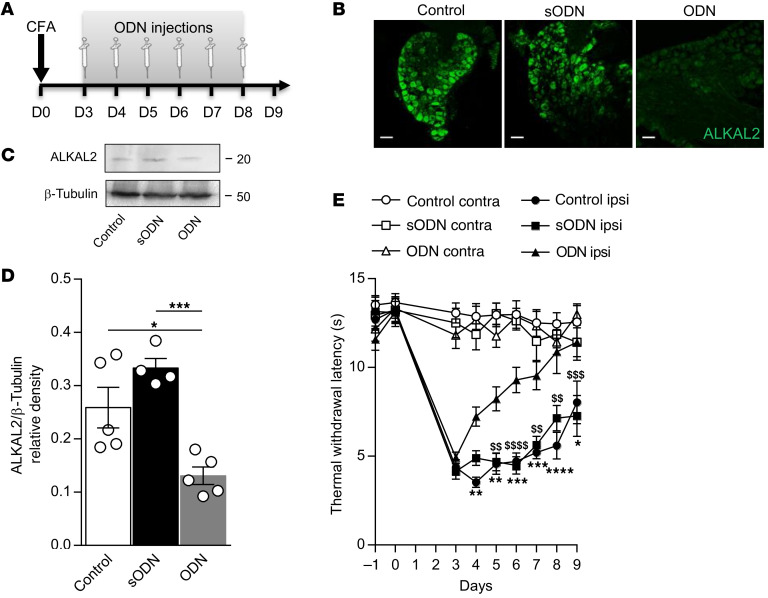
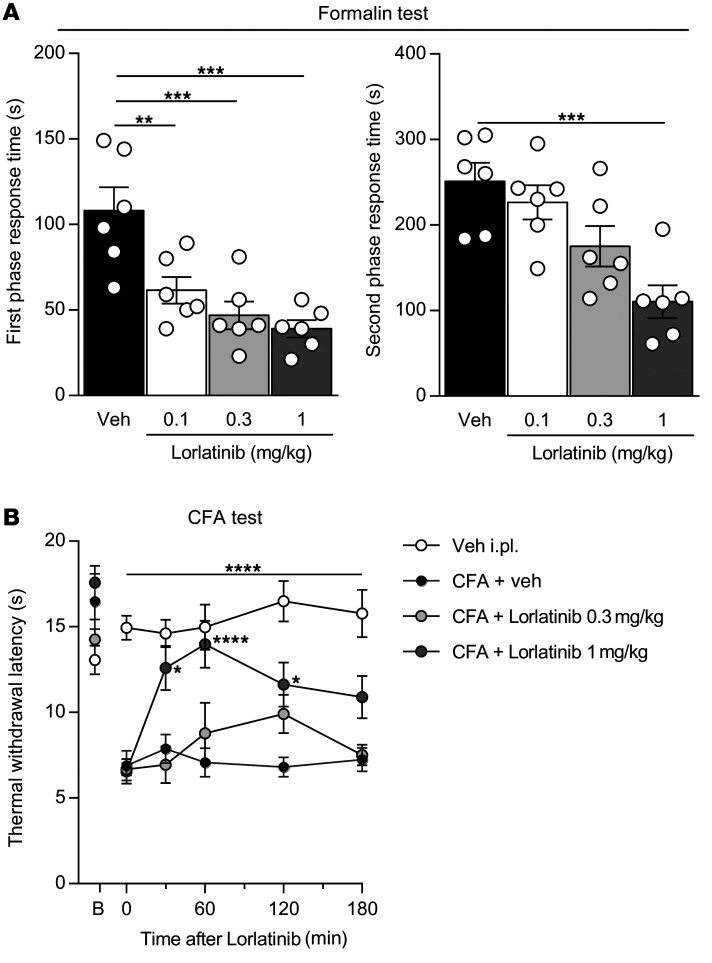
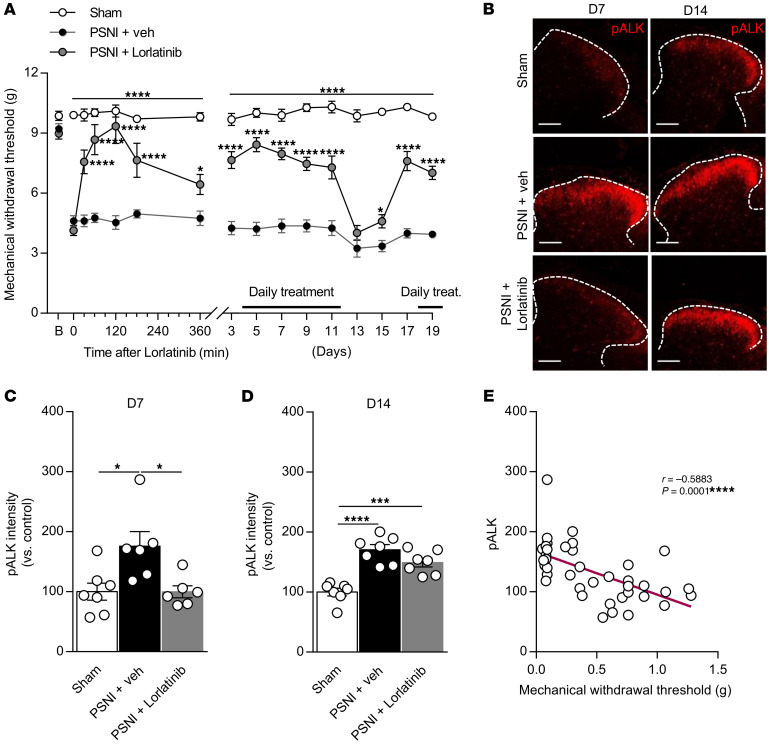
The anaplastic lymphoma kinase (ALK) is a receptor tyrosine kinase known for its oncogenic potential that is involved in the development of the peripheral and central nervous system. ALK receptor ligands ALKAL1 and ALKAL2 were recently found to promote neuronal differentiation and survival. Here, we show that inflammation or injury enhanced ALKAL2 expression in a subset of TRPV1+ sensory neurons. Notably, ALKAL2 was particularly enriched in both mouse and human peptidergic nociceptors, yet weakly expressed in nonpeptidergic, large-diameter myelinated neurons or in the brain. Using a coculture expression system, we found that nociceptors exposed to ALKAL2 exhibited heightened excitability and neurite outgrowth. Intraplantar CFA or intrathecal infusion of recombinant ALKAL2 led to ALK phosphorylation in the lumbar dorsal horn of the spinal cord. Finally, depletion of ALKAL2 in dorsal root ganglia or blocking ALK with clinically available compounds crizotinib or lorlatinib reversed thermal hyperalgesia and mechanical allodynia induced by inflammation or nerve injury, respectively. Overall, our work uncovers the ALKAL2/ALK signaling axis as a central regulator of nociceptor-induced sensitization. We propose that clinically approved ALK inhibitors used for non-small cell lung cancer and neuroblastomas could be repurposed to treat persistent pain conditions.
Keywords: Cancer; Neuroscience; Pain; Signal transduction.
Figures


References
-
- Coleridge JC, Coleridge HM. Afferent vagal C fibre innervation of the lungs and airways and its functional significance. Rev Physiol Biochem Pharmacol. 1984;99:1–110. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
