

Natural behavior is the language of the brain
- PMID: 35609550
- PMCID: PMC10082559
- DOI: 10.1016/j.cub.2022.03.031
Natural behavior is the language of the brain
Abstract
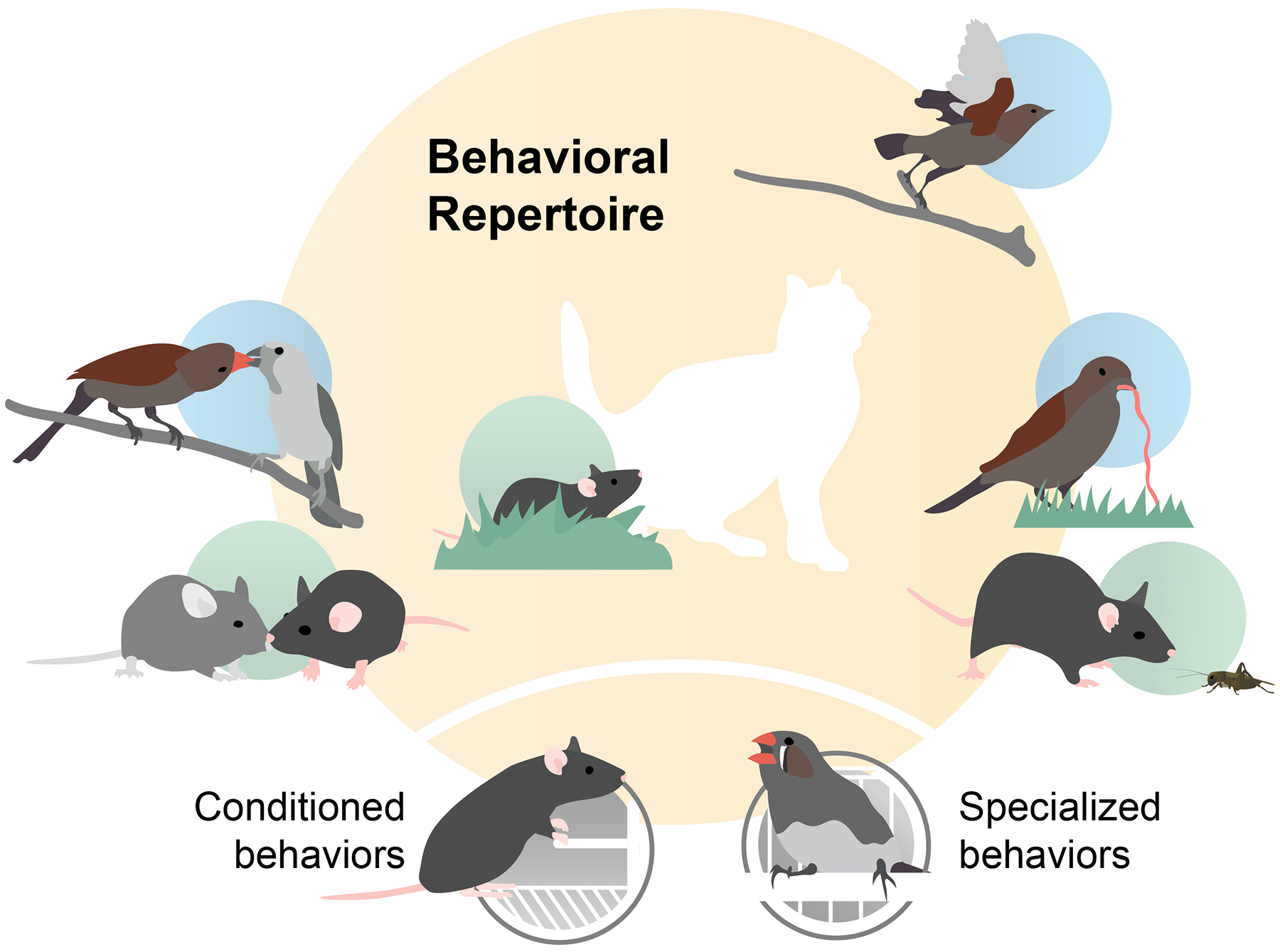
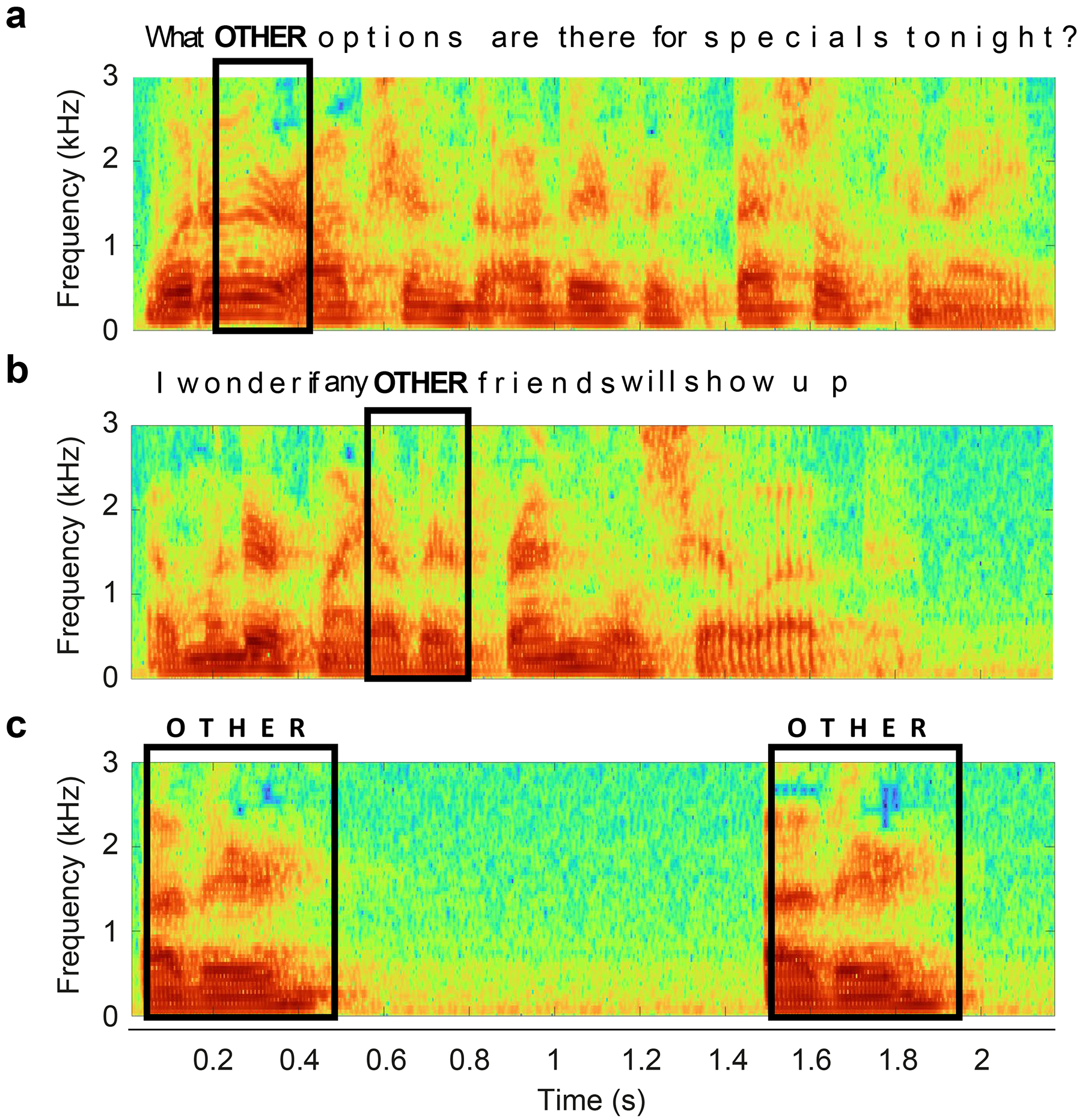
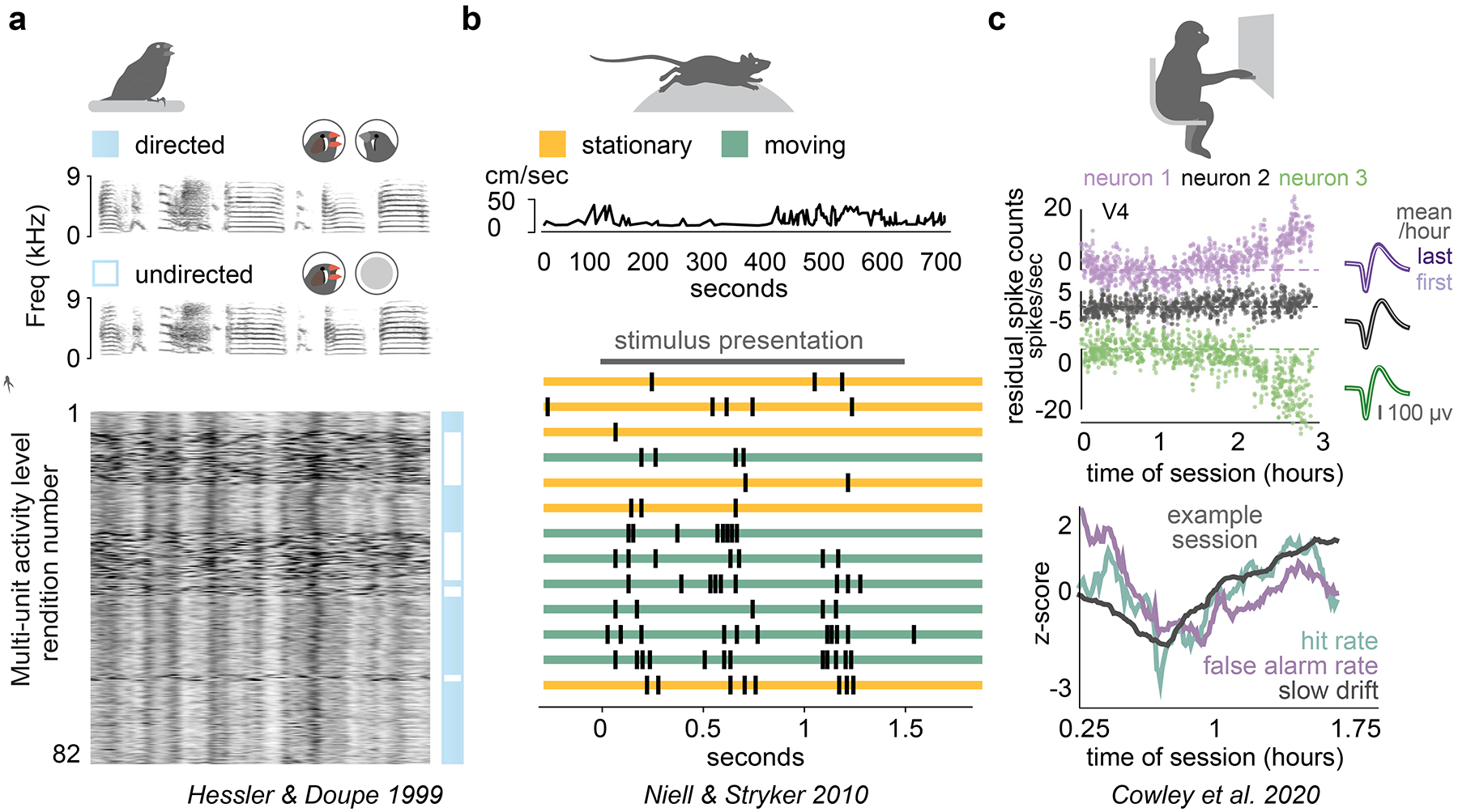
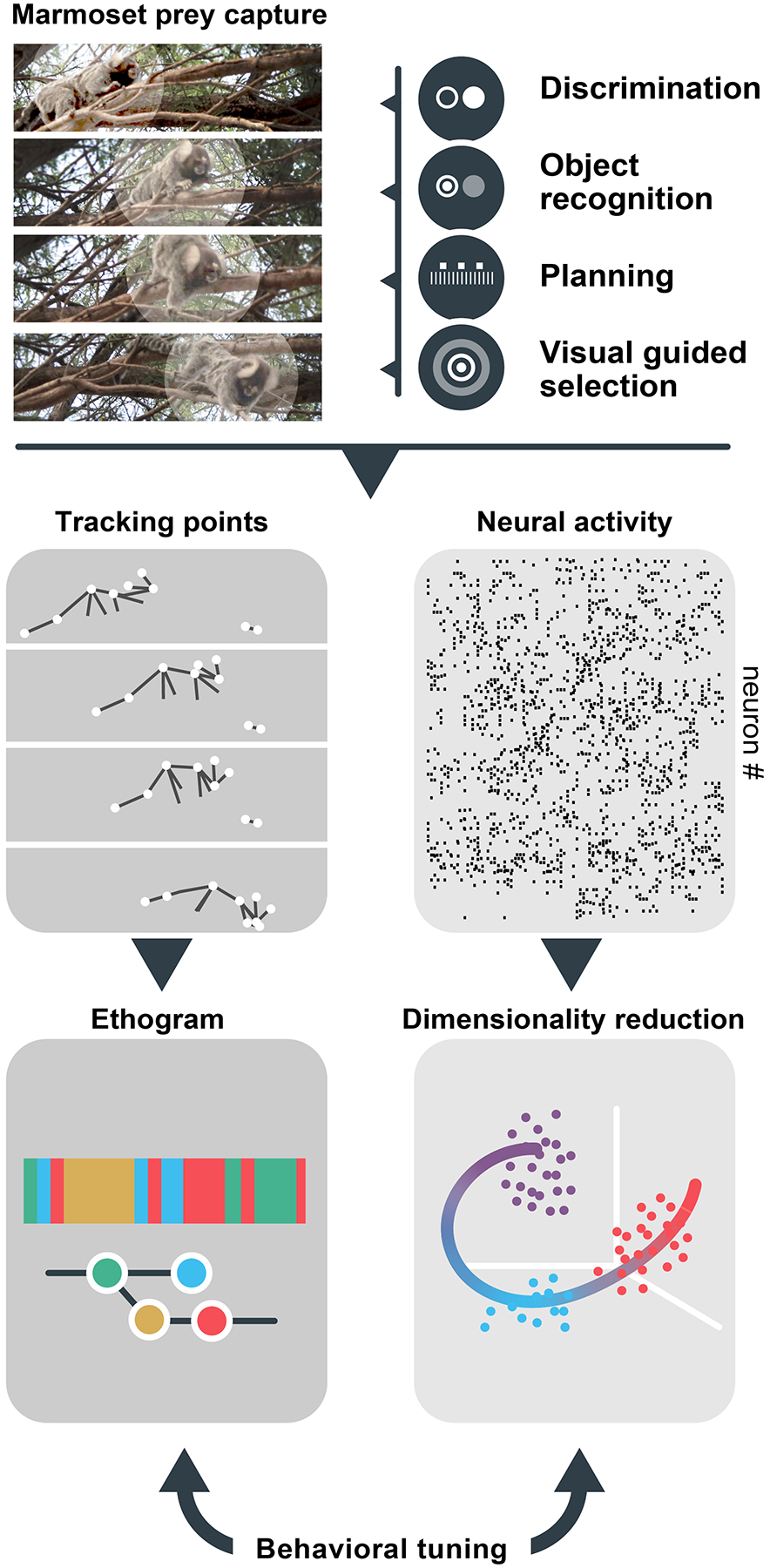
The breadth and complexity of natural behaviors inspires awe. Understanding how our perceptions, actions, and internal thoughts arise from evolved circuits in the brain has motivated neuroscientists for generations. Researchers have traditionally approached this question by focusing on stereotyped behaviors, either natural or trained, in a limited number of model species. This approach has allowed for the isolation and systematic study of specific brain operations, which has greatly advanced our understanding of the circuits involved. At the same time, the emphasis on experimental reductionism has left most aspects of the natural behaviors that have shaped the evolution of the brain largely unexplored. However, emerging technologies and analytical tools make it possible to comprehensively link natural behaviors to neural activity across a broad range of ethological contexts and timescales, heralding new modes of neuroscience focused on natural behaviors. Here we describe a three-part roadmap that aims to leverage the wealth of behaviors in their naturally occurring distributions, linking their variance with that of underlying neural processes to understand how the brain is able to successfully navigate the everyday challenges of animals' social and ecological landscapes. To achieve this aim, experimenters must harness one challenge faced by all neurobiological systems, namely variability, in order to gain new insights into the language of the brain.
Copyright © 2022 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
References
-
- Krakauer JW, Ghazanfar AA, Gomez-Marin A, MacIver MA, and Poeppel D (2017). Neuroscience needs behavior: correcting a reductionist bias. Neuron 93, 480–490. - PubMed
