

Dysfunctional dendritic cells limit antigen-specific T cell response in glioma
- PMID: 35609569
- PMCID: PMC9925697
- DOI: 10.1093/neuonc/noac138
Dysfunctional dendritic cells limit antigen-specific T cell response in glioma
Abstract
Background: Dendritic cells (DC), the most potent professional antigen presenting cells capable of effective cross-presentation, have been demonstrated to license T helper cells to induce antitumor immunity in solid tumors. Specific DC subtypes are recruited to the injured brain by microglial chemokines, locally adapting to distinct transcriptional profiles. In isocitrate dehydrogenase (IDH) type 1 mutant gliomas, monocyte-derived macrophages have recently been shown to display an attenuated intratumoral antigen presentation capacity as consequence of the local accumulation of the oncometabolite R-2-hydroxyglutarate. The functionality and the contribution of DC to the IDH-mutant tumor microenvironment (TME) remains unclear.
Methods: Frequencies and intratumoral phenotypes of human DC in IDH-wildtype (IDHwt) and -mutant high-grade gliomas are comparatively assessed by transcriptomic and proteomic profiling. DC functionality is investigated in experimental murine glioblastomas expressing the model antigen ovalbumin. Single-cell sequencing-based pseudotime analyses and spectral flow cytometric analyses are used to profile DC states longitudinally.
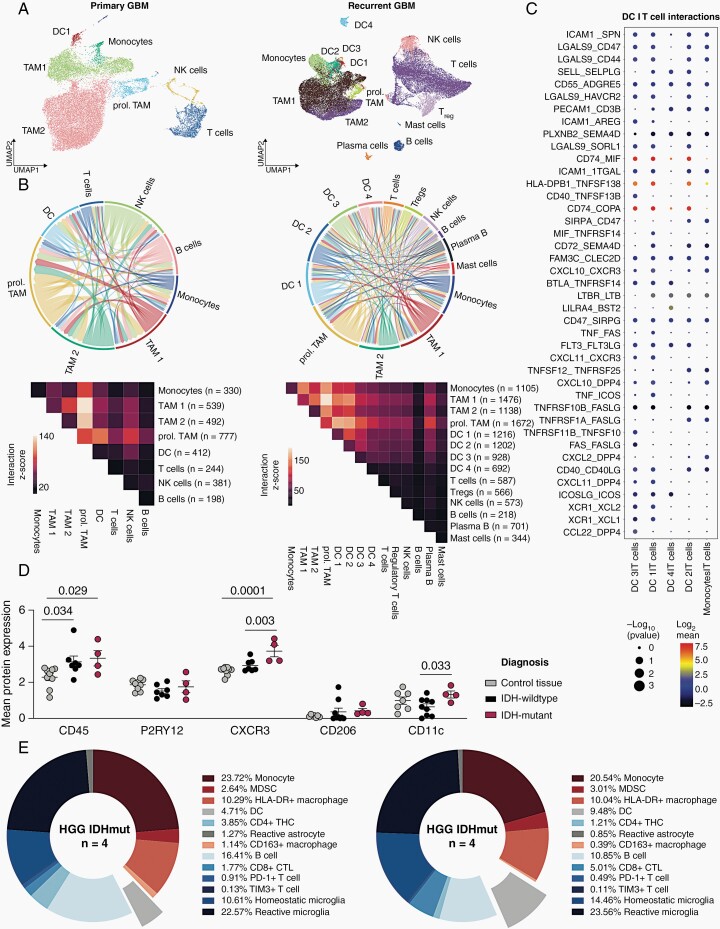
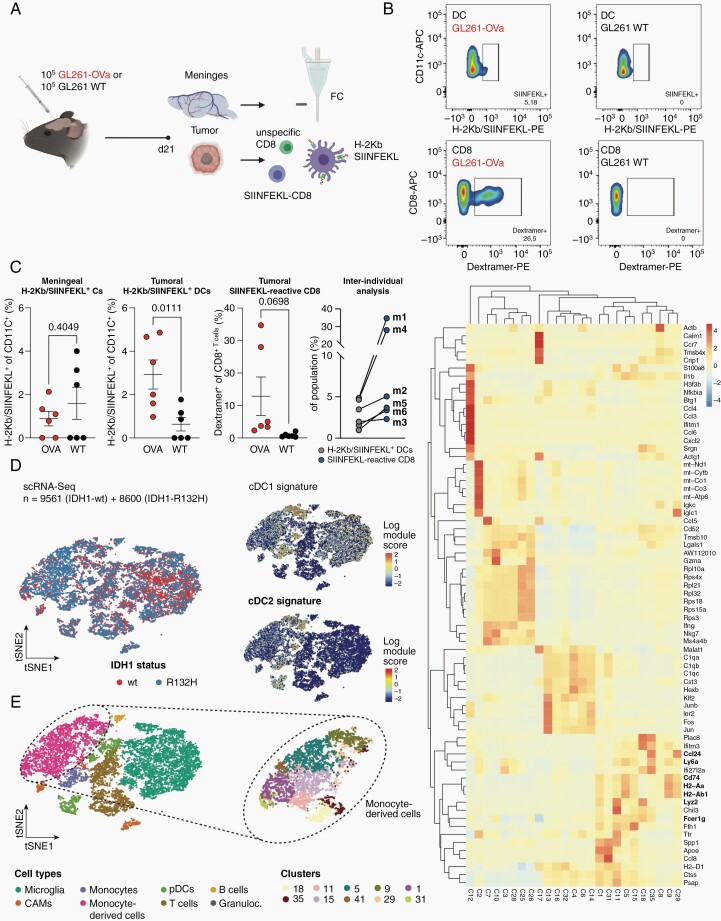
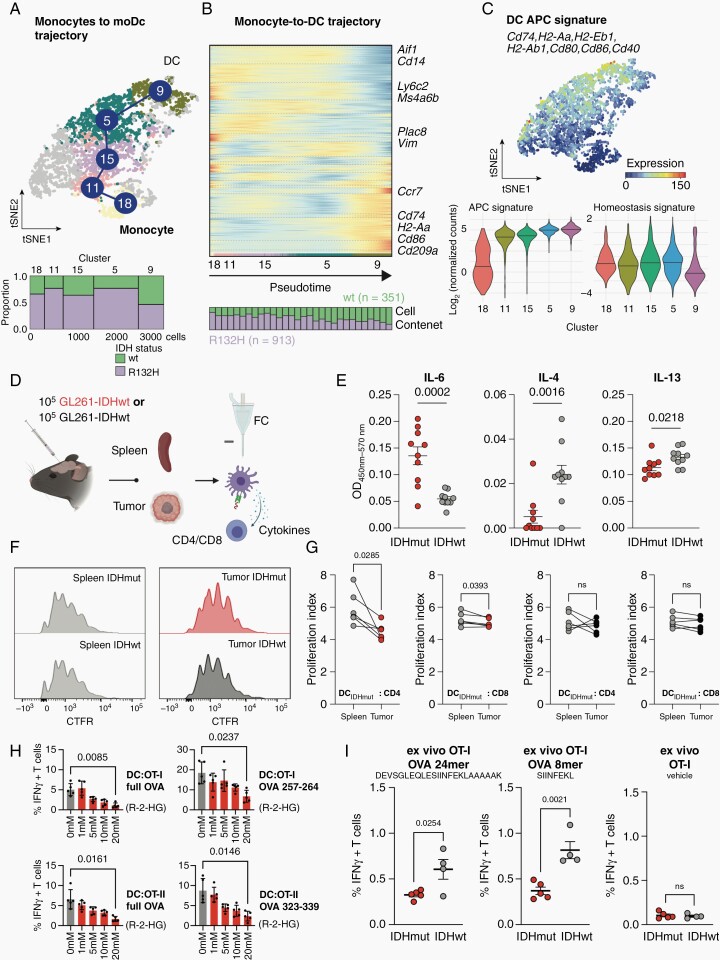
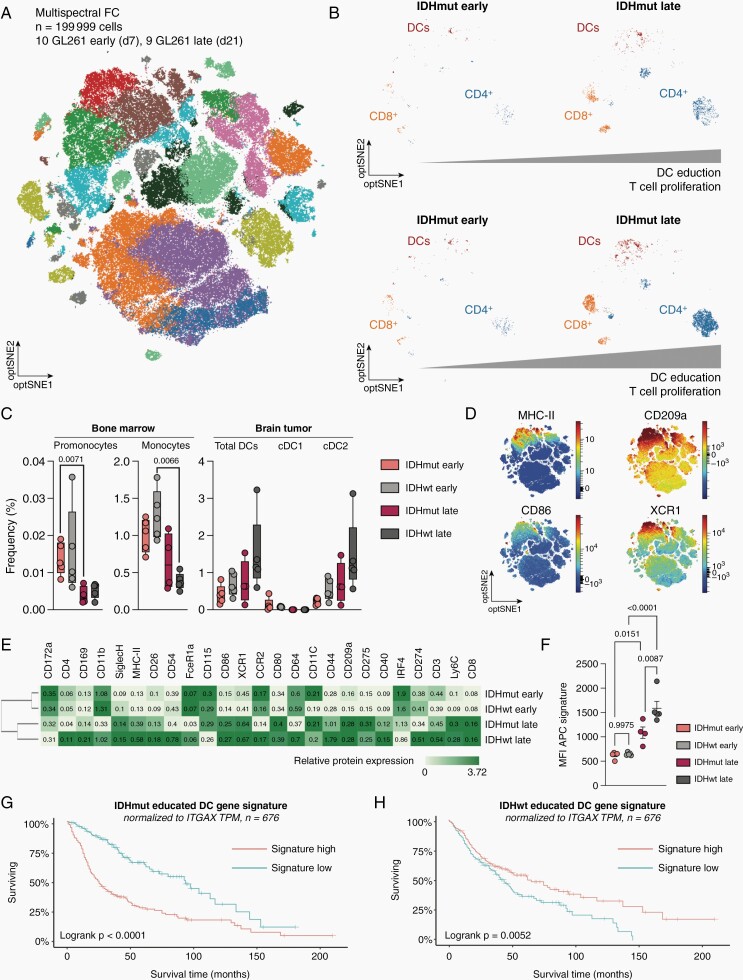
Results: DC are present in primary and recurrent high-grade gliomas and interact with other immune cell types within the TME. In murine glioblastomas, we find an IDH-status-associated major histocompatibility class I-restricted cross-presentation of tumor antigens by DC specifically in the tumor but not in meninges or secondary lymphoid organs of tumor-bearing animals. In single-cell sequencing-based pseudotime and longitudinal spectral flow cytometric analyses, we demonstrate an IDH-status-dependent differential, exclusively microenvironmental education of DC.
Conclusions: Glioma-associated DCs are relevantly abundant in human IDHwt and mutant tumors. Glioma IDH mutations result in specifically educated, dysfunctional DCs via paracrine reprogramming of infiltrating monocytes, providing the basis for combinatorial immunotherapy concepts against IDH mutant gliomas.
Keywords: IDH mutation; R-2-HG; cDC1; cDC2; dendritic cell; glioblastoma; glioma microenvironment.
© The Author(s) 2022. Published by Oxford University Press on behalf of the Society for Neuro-Oncology.
Figures
Comment in
-
Commentary on "Dysfunctional dendritic cells limit antigen-specific T cell response in glioma".Neuro Oncol. 2023 Feb 14;25(2):277-278. doi: 10.1093/neuonc/noac256. Neuro Oncol. 2023. PMID: 36445342 Free PMC article. No abstract available.
References
-
- Omuro A, DeAngelis LM. Glioblastoma and other malignant gliomas: a clinical review. J Am Med Assoc. 2013; 310(17):1842–1850. - PubMed
-
- Friebel E, Kapolou K, Unger S, et al. Single-cell mapping of human brain cancer reveals tumor-specific instruction of tissue-invading leukocytes. Cell. 2020; 181(17):1626–1642.e20. - PubMed
-
- Locarno C V., Simonelli M, Carenza C, et al. Role of myeloid cells in the immunosuppressive microenvironment in gliomas. Immunobiology. 2020; 225(1):151853. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
