

Defining mitochondrial protein functions through deep multiomic profiling
- PMID: 35614220
- PMCID: PMC9310563
- DOI: 10.1038/s41586-022-04765-3
Defining mitochondrial protein functions through deep multiomic profiling
Abstract
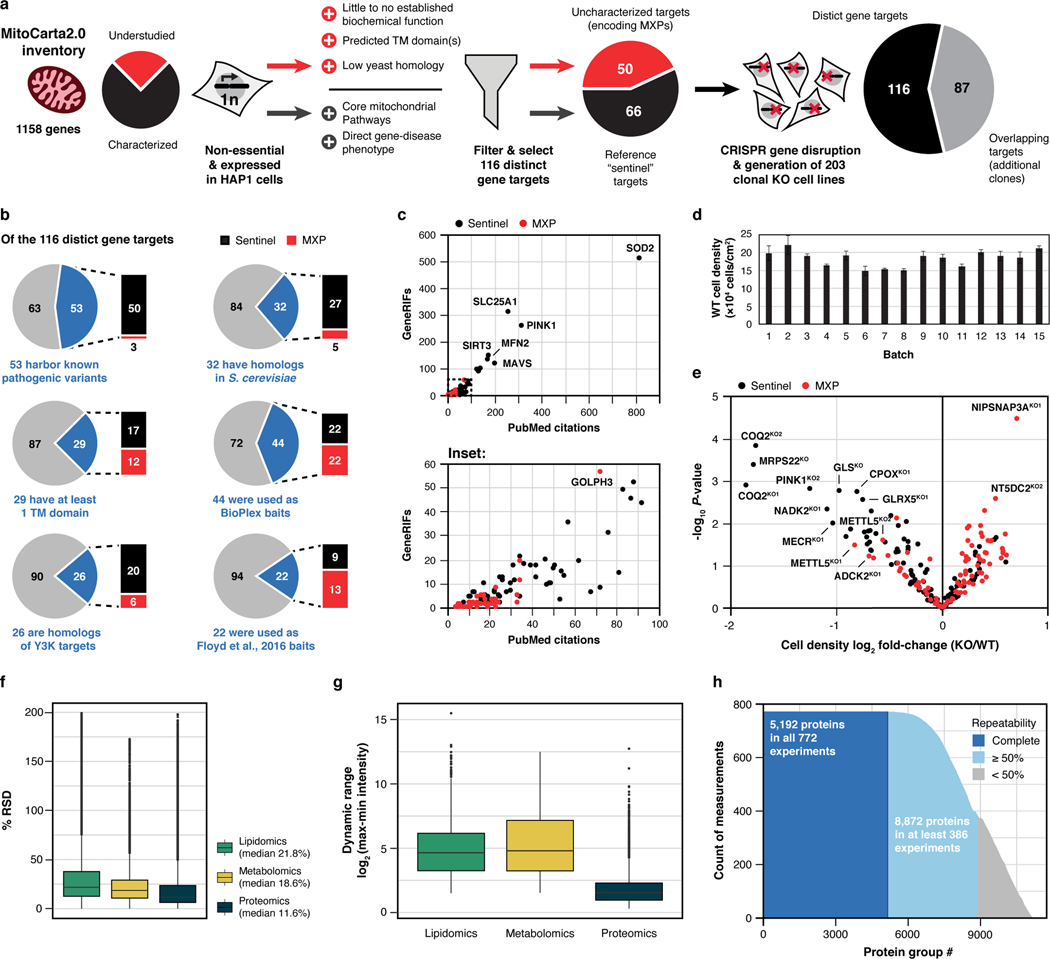
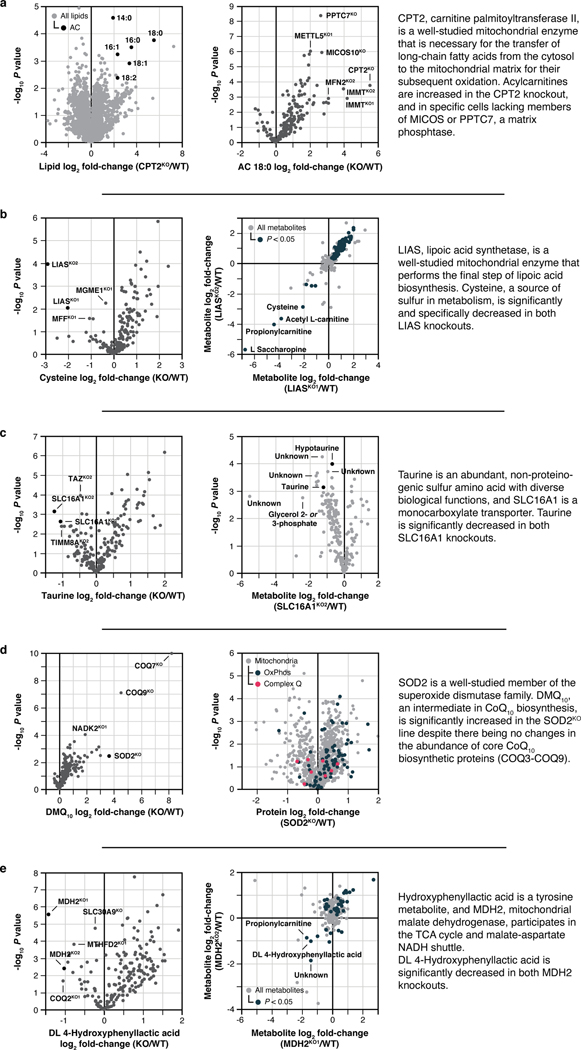
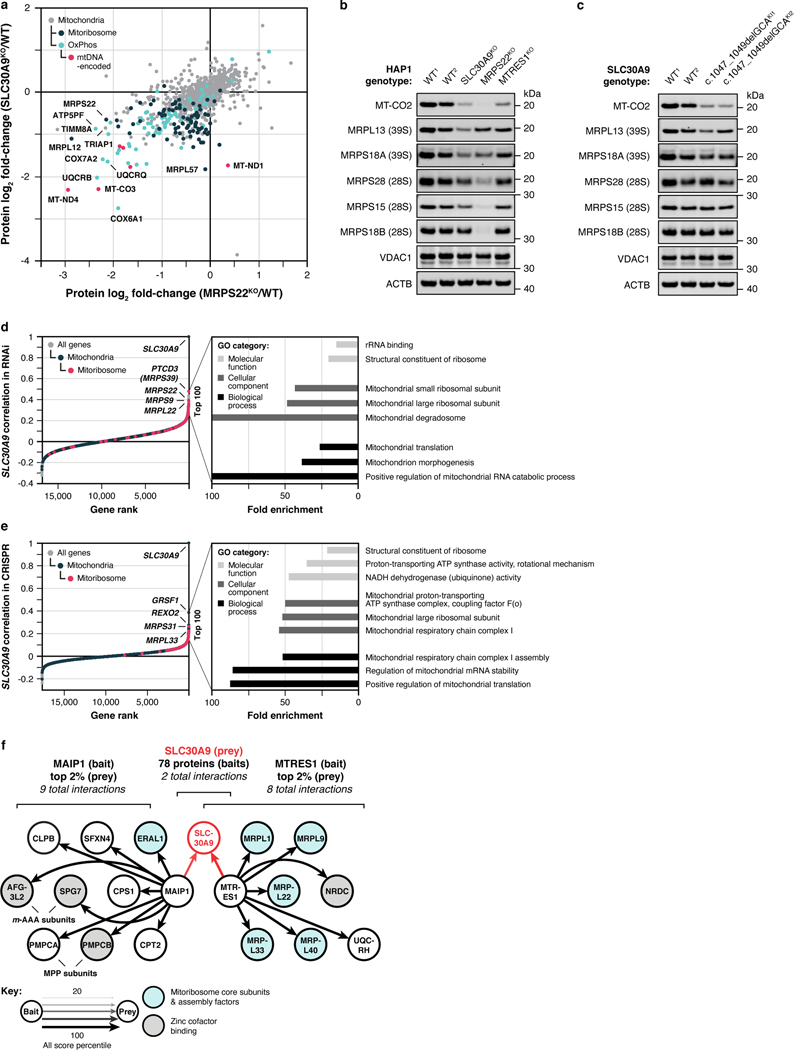
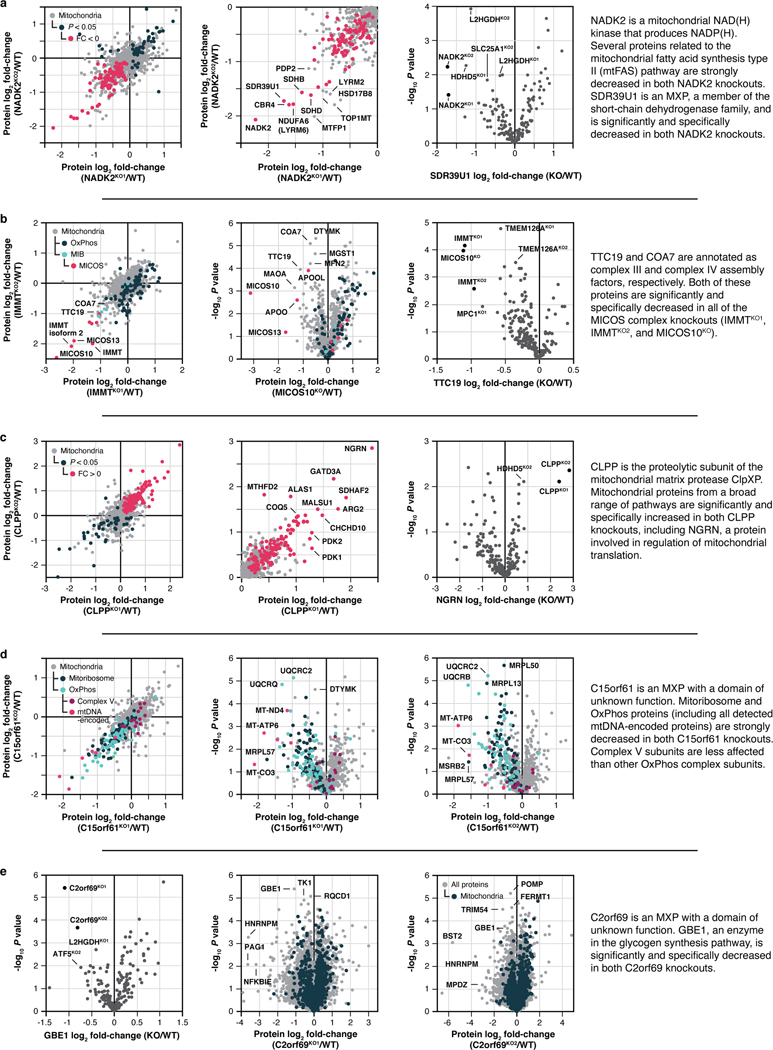
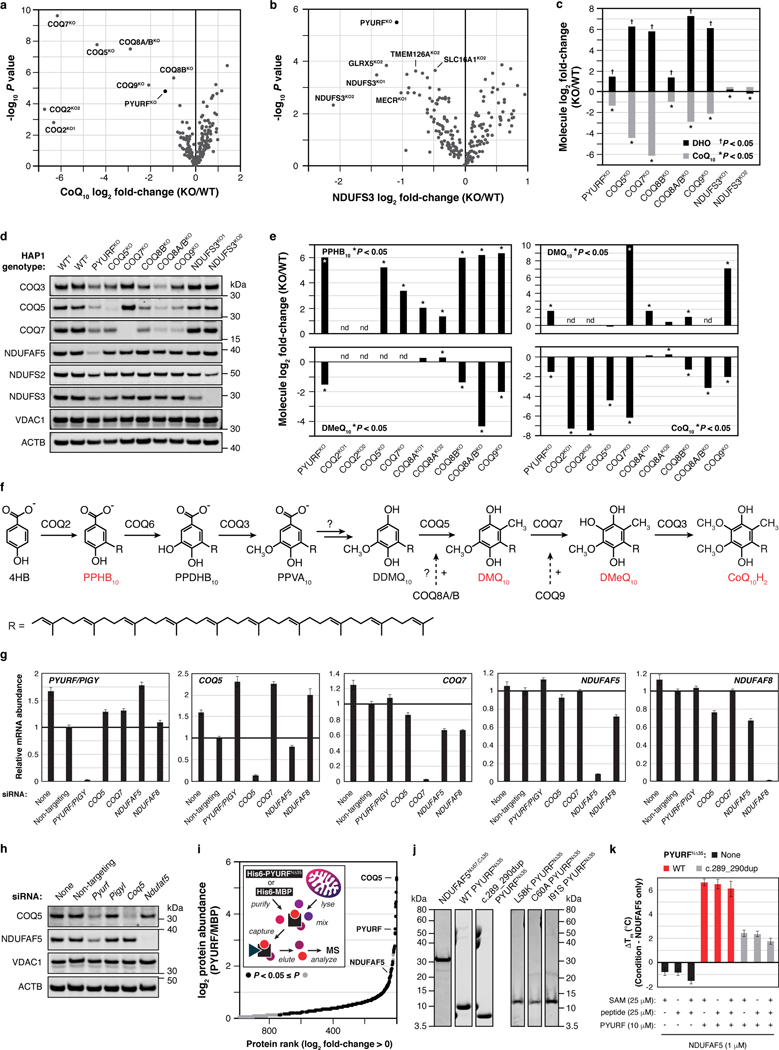
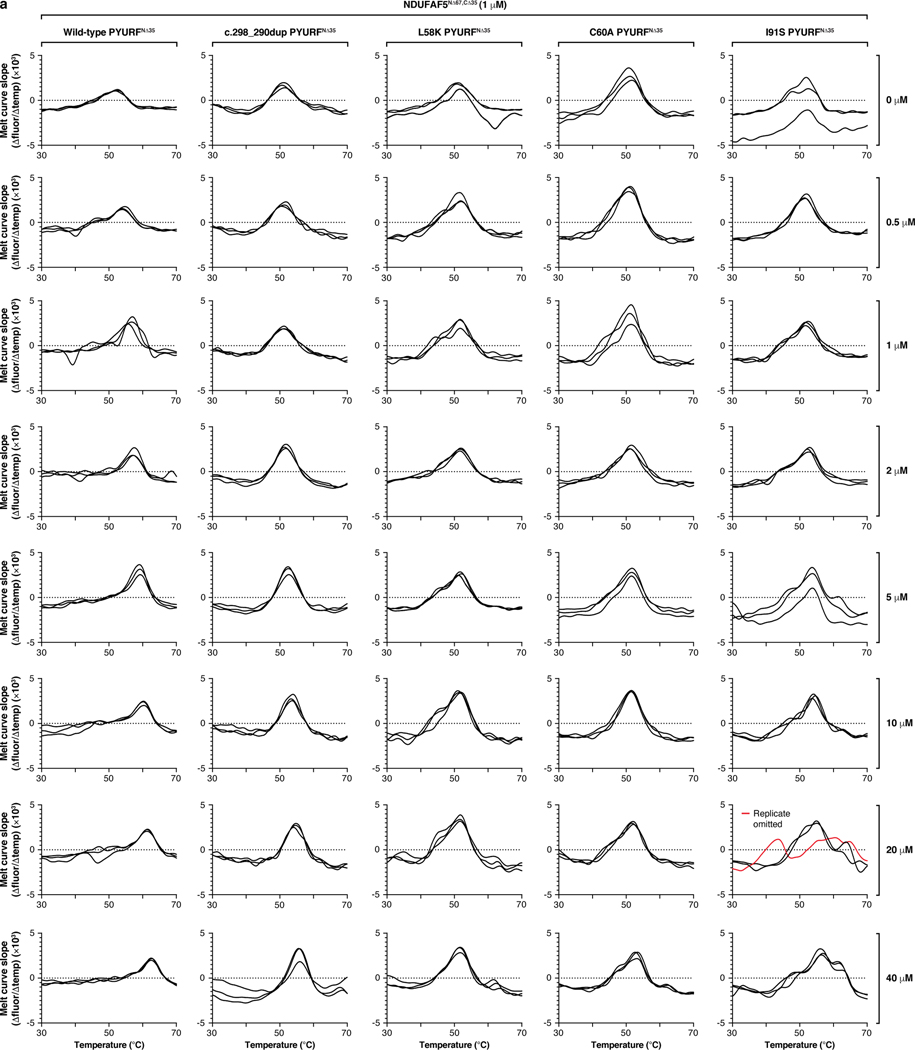
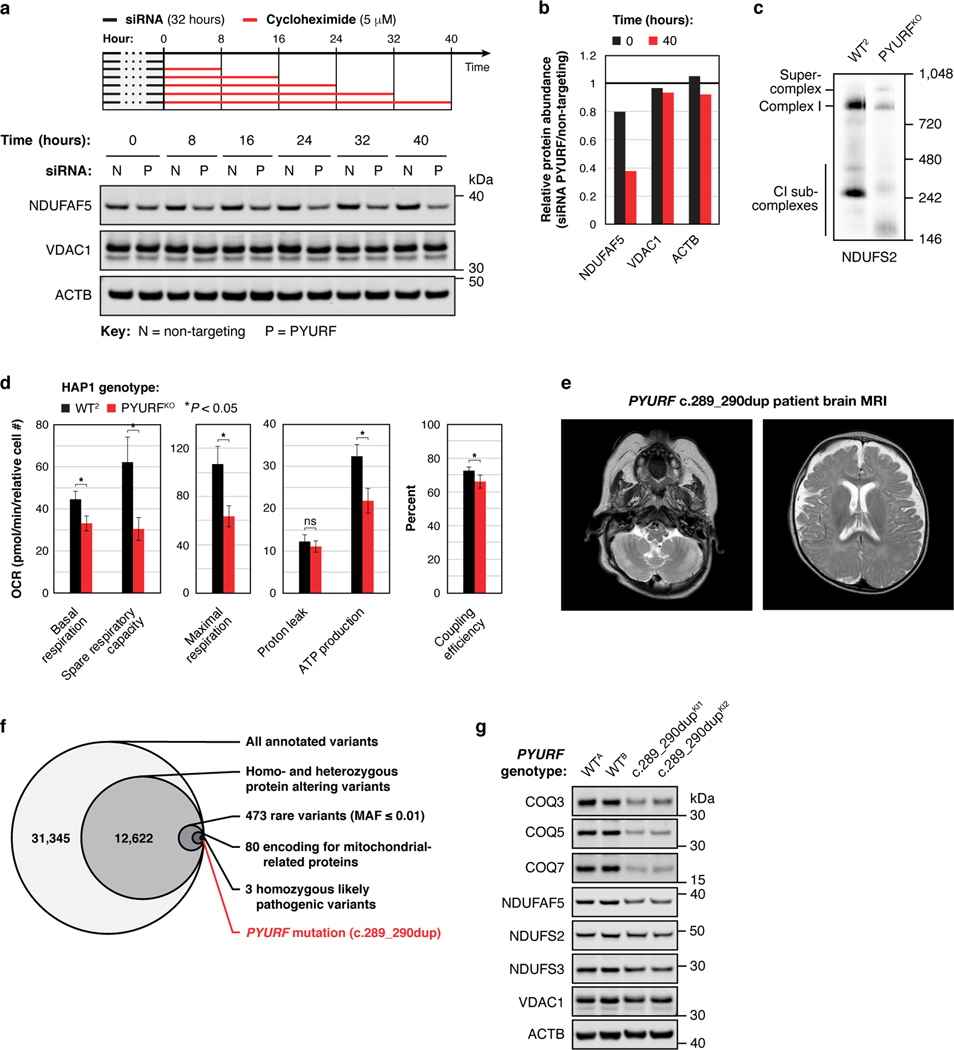
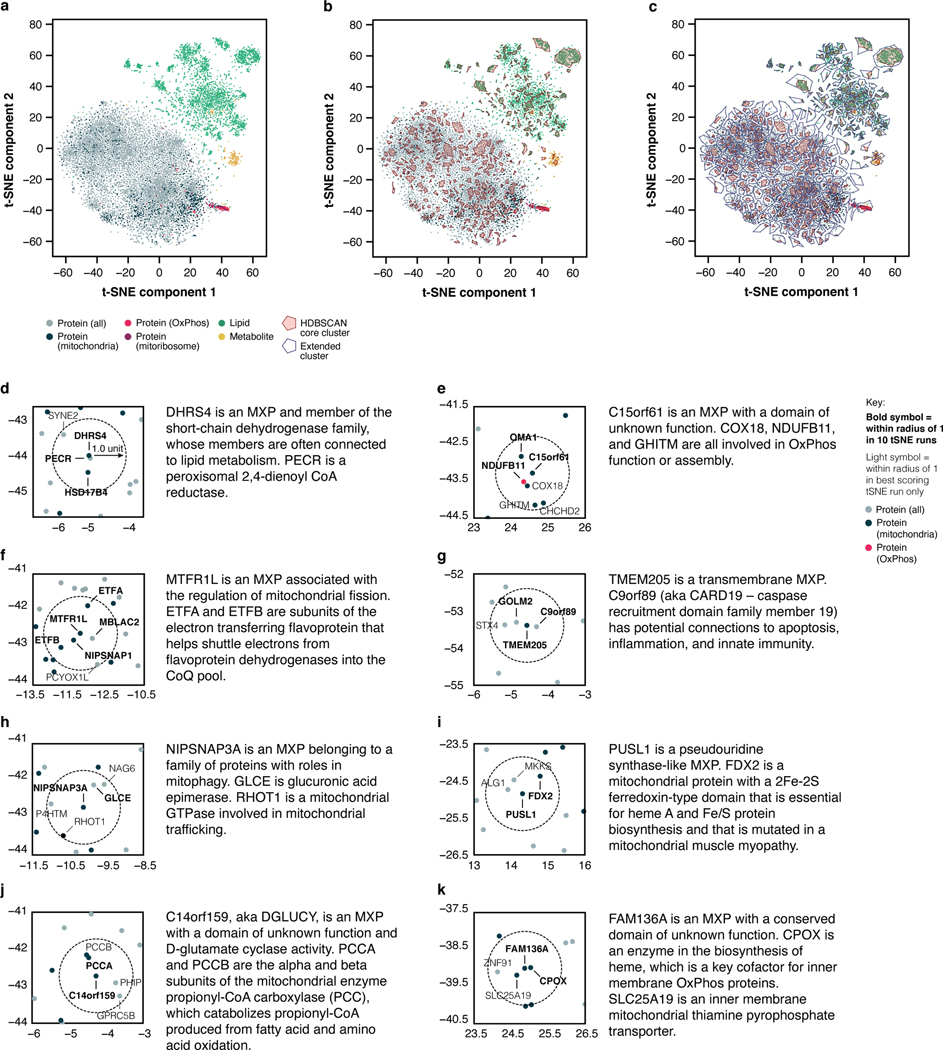
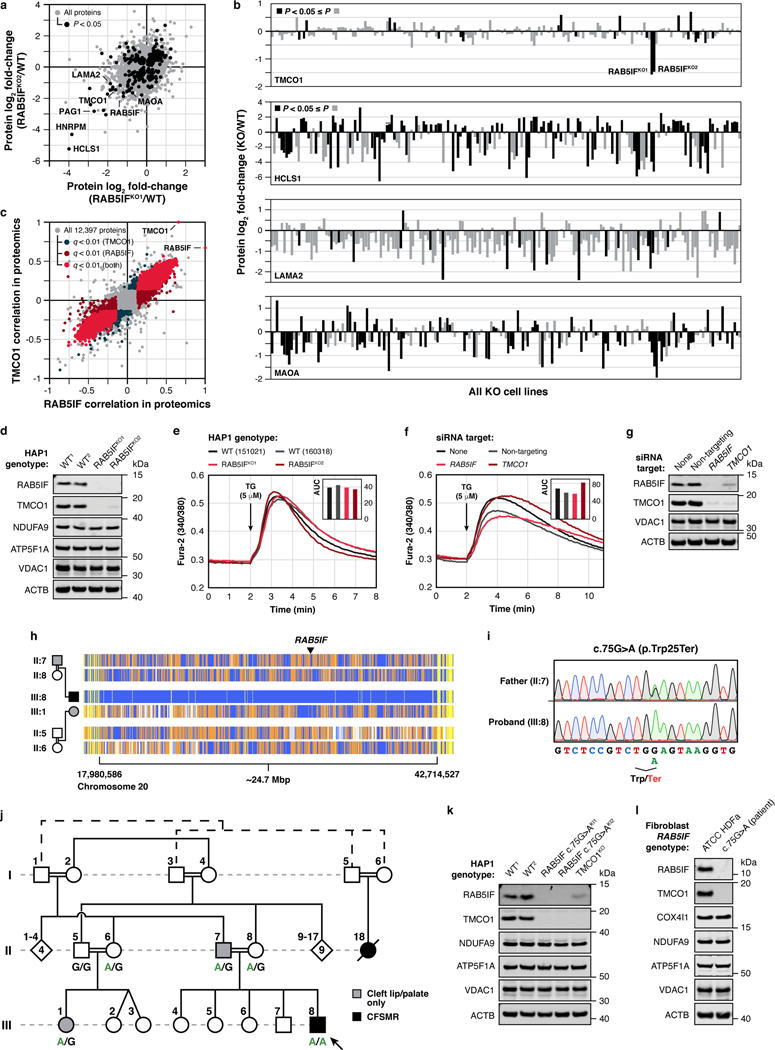
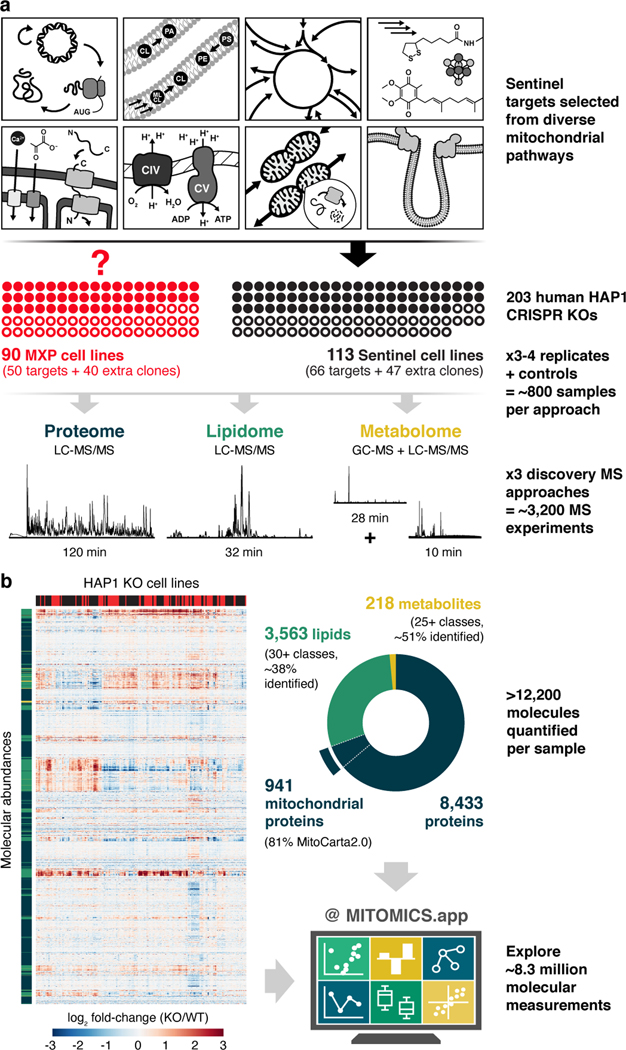
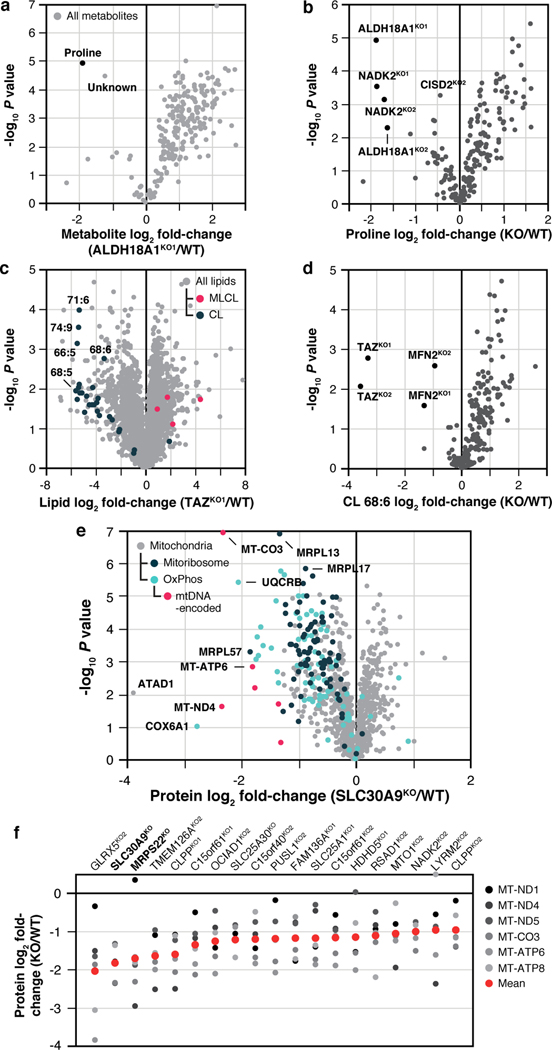
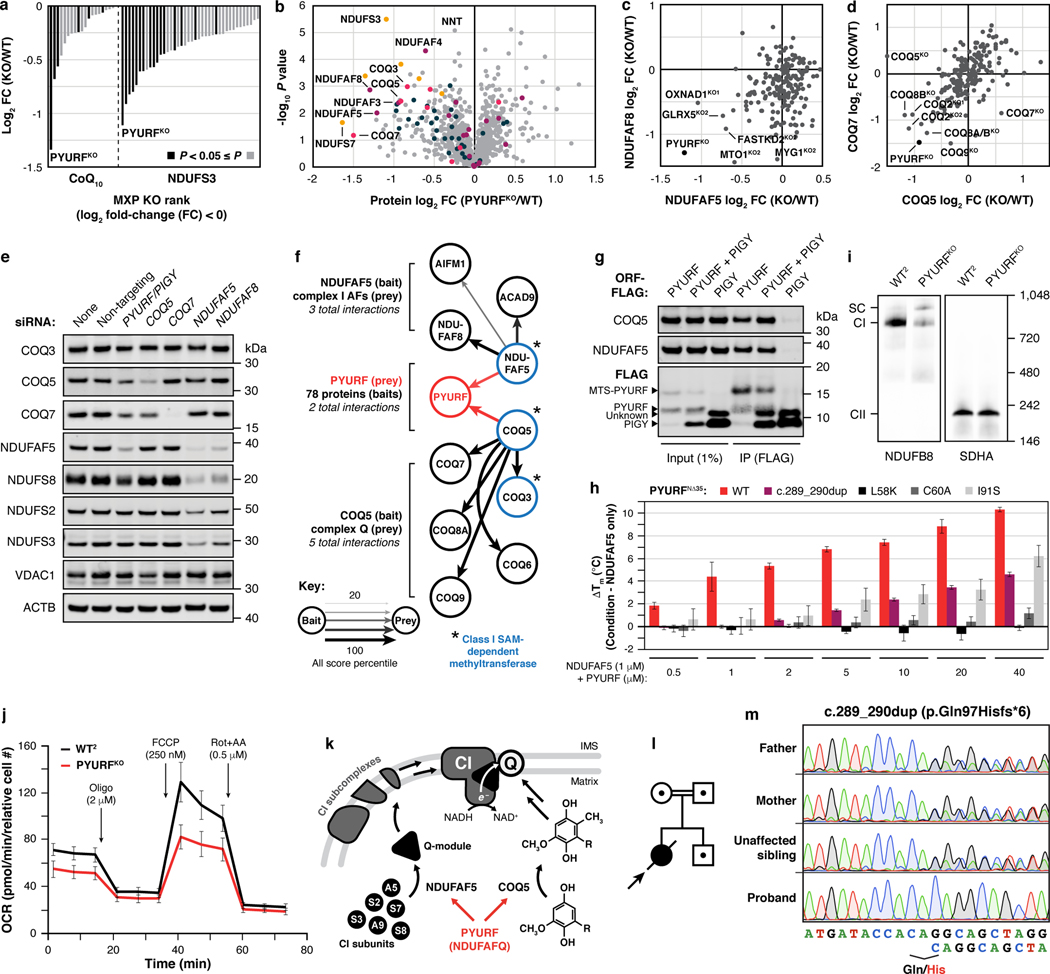
Mitochondria are epicentres of eukaryotic metabolism and bioenergetics. Pioneering efforts in recent decades have established the core protein componentry of these organelles1 and have linked their dysfunction to more than 150 distinct disorders2,3. Still, hundreds of mitochondrial proteins lack clear functions4, and the underlying genetic basis for approximately 40% of mitochondrial disorders remains unresolved5. Here, to establish a more complete functional compendium of human mitochondrial proteins, we profiled more than 200 CRISPR-mediated HAP1 cell knockout lines using mass spectrometry-based multiomics analyses. This effort generated approximately 8.3 million distinct biomolecule measurements, providing a deep survey of the cellular responses to mitochondrial perturbations and laying a foundation for mechanistic investigations into protein function. Guided by these data, we discovered that PIGY upstream open reading frame (PYURF) is an S-adenosylmethionine-dependent methyltransferase chaperone that supports both complex I assembly and coenzyme Q biosynthesis and is disrupted in a previously unresolved multisystemic mitochondrial disorder. We further linked the putative zinc transporter SLC30A9 to mitochondrial ribosomes and OxPhos integrity and established RAB5IF as the second gene harbouring pathogenic variants that cause cerebrofaciothoracic dysplasia. Our data, which can be explored through the interactive online MITOMICS.app resource, suggest biological roles for many other orphan mitochondrial proteins that still lack robust functional characterization and define a rich cell signature of mitochondrial dysfunction that can support the genetic diagnosis of mitochondrial diseases.
© 2022. The Author(s), under exclusive licence to Springer Nature Limited.
Conflict of interest statement
Competing interest declaration
J.J.C. is a consultant for Thermo Fisher Scientific.
Figures
Comment in
-
Mining the mitochondrial proteome.Nat Rev Mol Cell Biol. 2022 Jul;23(7):447. doi: 10.1038/s41580-022-00503-9. Nat Rev Mol Cell Biol. 2022. PMID: 35676505 No abstract available.
-
Systems biochemistry to "deorphanize" human mitochondrial proteome.Mol Cell. 2022 Aug 4;82(15):2735-2737. doi: 10.1016/j.molcel.2022.07.005. Mol Cell. 2022. PMID: 35931038
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
