

Investigating Abiotic and Biotic Mechanisms of Pyrite Reduction
- PMID: 35615515
- PMCID: PMC9124975
- DOI: 10.3389/fmicb.2022.878387
Investigating Abiotic and Biotic Mechanisms of Pyrite Reduction
Abstract
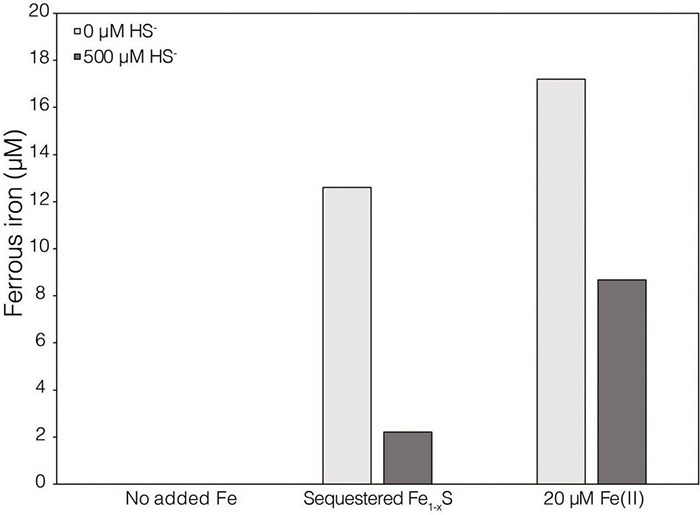
Pyrite (FeS2) has a very low solubility and therefore has historically been considered a sink for iron (Fe) and sulfur (S) and unavailable to biology in the absence of oxygen and oxidative weathering. Anaerobic methanogens were recently shown to reduce FeS2 and assimilate Fe and S reduction products to meet nutrient demands. However, the mechanism of FeS2 mineral reduction and the forms of Fe and S assimilated by methanogens remained unclear. Thermodynamic calculations described herein indicate that H2 at aqueous concentrations as low as 10-10 M favors the reduction of FeS2, with sulfide (HS-) and pyrrhotite (Fe1- x S) as products; abiotic laboratory experiments confirmed the reduction of FeS2 with dissolved H2 concentrations greater than 1.98 × 10-4 M H2. Growth studies of Methanosarcina barkeri provided with FeS2 as the sole source of Fe and S resulted in H2 production but at concentrations too low to drive abiotic FeS2 reduction, based on abiotic laboratory experimental data. A strain of M. barkeri with deletions in all [NiFe]-hydrogenases maintained the ability to reduce FeS2 during growth, providing further evidence that extracellular electron transport (EET) to FeS2 does not involve H2 or [NiFe]-hydrogenases. Physical contact between cells and FeS2 was required for mineral reduction but was not required to obtain Fe and S from dissolution products. The addition of a synthetic electron shuttle, anthraquinone-2,6-disulfonate, allowed for biological reduction of FeS2 when physical contact between cells and FeS2 was prohibited, indicating that exogenous electron shuttles can mediate FeS2 reduction. Transcriptomics experiments revealed upregulation of several cytoplasmic oxidoreductases during growth of M. barkeri on FeS2, which may indicate involvement in provisioning low potential electrons for EET to FeS2. Collectively, the data presented herein indicate that reduction of insoluble FeS2 by M. barkeri occurred via electron transfer from the cell surface to the mineral surface resulting in the generation of soluble HS- and mineral-associated Fe1- x S. Solubilized Fe(II), but not HS-, from mineral-associated Fe1- x S reacts with aqueous HS- yielding aqueous iron sulfur clusters (FeS aq ) that likely serve as the Fe and S source for methanogen growth and activity. FeS aq nucleation and subsequent precipitation on the surface of cells may result in accelerated EET to FeS2, resulting in positive feedback between cell activity and FeS2 reduction.
Keywords: dissolution; extracellular electron transfer; hydrogen; methanogens; pyrite (FeS2); pyrrhotite (Fe1–xS).
Copyright © 2022 Spietz, Payne, Kulkarni, Metcalf, Roden and Boyd.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures








References
-
- Beifuss U., Tietze M. (2005). “Methanophenazine and other natural biologically active phenazines,” in Natural Products Synthesis II. Topics in Current Chemistry, ed. Mulzer J. (Berlin: Springer; ), 77–113. 10.1007/b96889 - DOI
-
- Benning L. G., Wilkin R. T., Barnes H. L. (2000). Reaction pathways in the Fe-S system below 100°C. Chem. Geol. 167 25–51. 10.1016/s0009-2541(99)00198-9 - DOI
-
- Berner R. A. (1967). Thermodynamic stability of sedimentary iron sulfides. Am. J. Sci. 265 773–785. 10.2475/ajs.265.9.773 - DOI
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous