

Skewed Cellular Distribution and Low Activation of Functional T-Cell Responses in SARS-CoV-2 Non-Seroconvertors
- PMID: 35619701
- PMCID: PMC9128381
- DOI: 10.3389/fimmu.2022.815041
Skewed Cellular Distribution and Low Activation of Functional T-Cell Responses in SARS-CoV-2 Non-Seroconvertors
Abstract
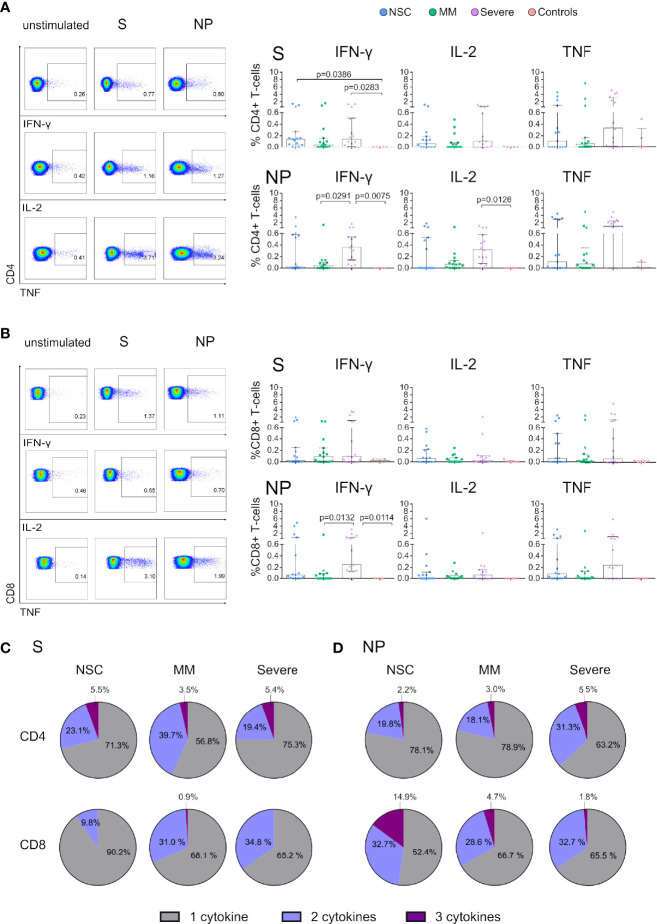
The role of T cells in the control of SARS-CoV-2 infection has been underestimated in favor of neutralizing antibodies. However, cellular immunity is essential for long-term viral control and protection from disease severity. To understand T-cell immunity in the absence of antibody generation we focused on a group of SARS-CoV-2 Non-Seroconvertors (NSC) recovered from infection. We performed an immune comparative analysis of SARS-CoV-2 infected individuals stratified by the absence or presence of seroconversion and disease severity. We report high levels of total naïve and low effector CD8+ T cells in NSC. Moreover, reduced levels of T-cell activation monitored by PD-1 and activation-induced markers were observed in the context of functional SARS-CoV-2 T-cell responses. Longitudinal data indicate the stability of the NSC phenotype over three months of follow-up after infection. Together, these data characterized distinctive immunological traits in NSC including skewed cellular distribution, low activation and functional SARS-CoV-2 T-cell responses. This data highlights the value of T-cell immune monitoring in populations with low seroconversion rates in response to SARS-CoV-2 infection and vaccination.
Keywords: SARS-CoV-2; T cell subsets; cellular immunity; function; immune activation; non-seroconvertor.
Copyright © 2022 Kilpeläinen, Jimenez-Moyano, Blanch-Lombarte, Ouchi, Peña, Quirant-Sanchez, Perez-Caballero, Chamorro, Blanco, Martínez-Caceres, Paredes, Mateu, Carrillo, Blanco, Brander, Massanella, Clotet and Prado.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures


References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous