

Altered synaptic plasticity of the longitudinal dentate gyrus network in noise-induced anxiety
- PMID: 35620435
- PMCID: PMC9127171
- DOI: 10.1016/j.isci.2022.104364
Altered synaptic plasticity of the longitudinal dentate gyrus network in noise-induced anxiety
Abstract
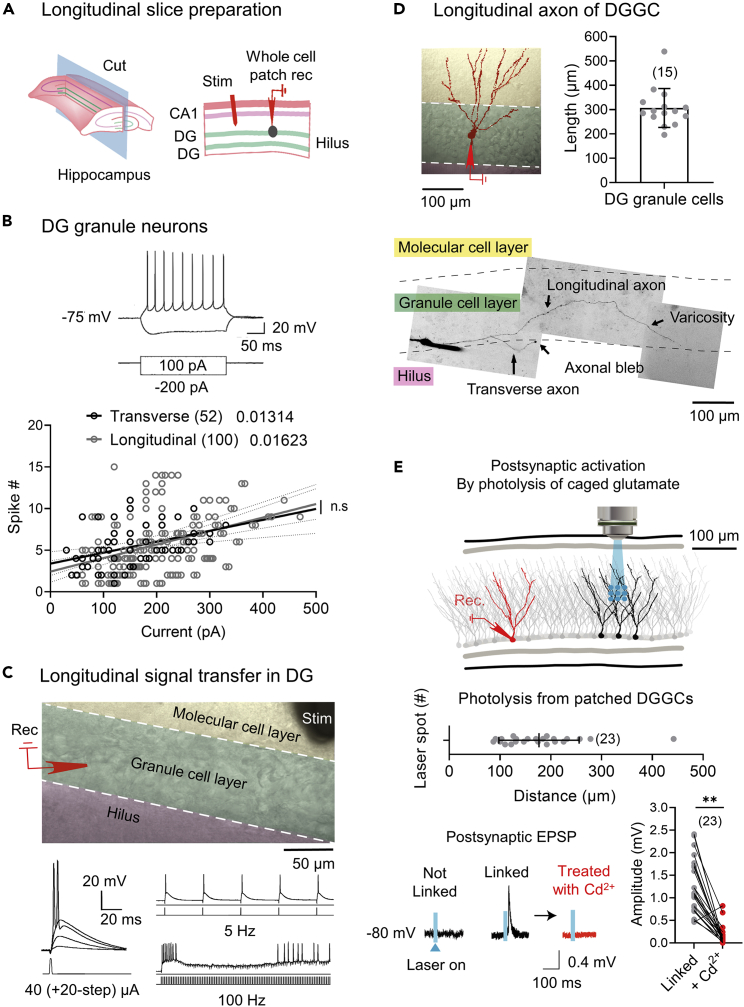
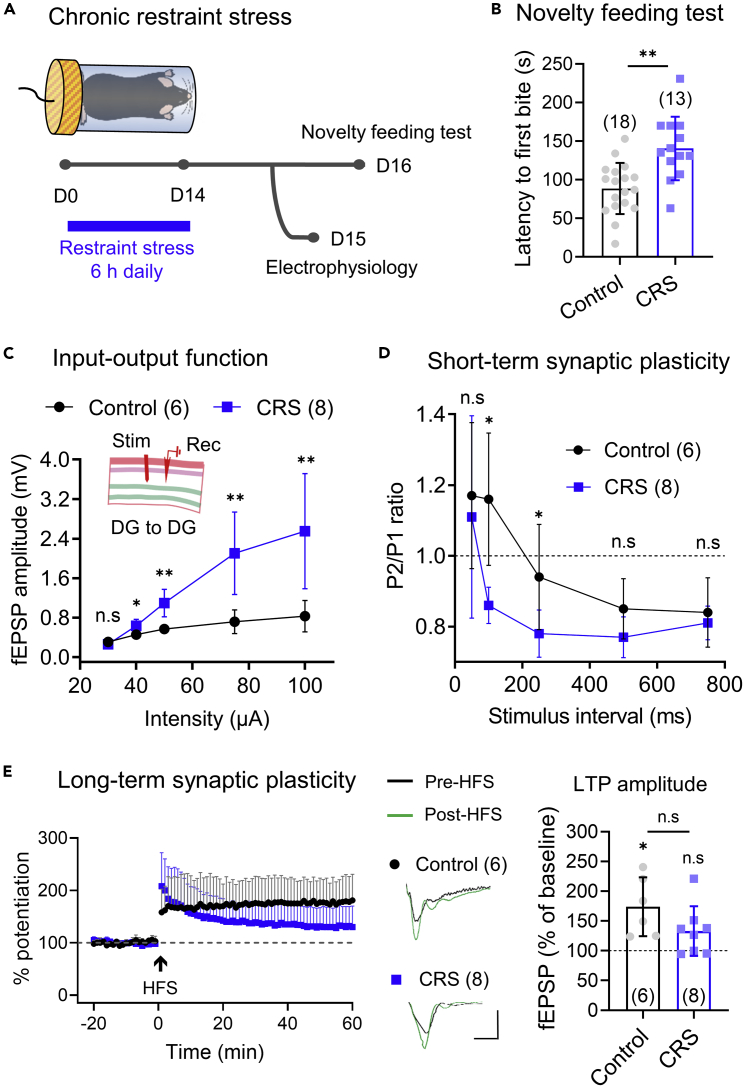
Anxiety is characteristic comorbidity of noise-induced hearing loss (NIHL), which causes physiological changes within the dentate gyrus (DG), a subfield of the hippocampus that modulates anxiety. However, which DG circuit underlies hearing loss-induced anxiety remains unknown. We utilize an NIHL mouse model to investigate short- and long-term synaptic plasticity in DG networks. The recently discovered longitudinal DG-DG network is a collateral of DG neurons synaptically connected with neighboring DG neurons and displays robust synaptic efficacy and plasticity. Furthermore, animals with NIHL demonstrate increased anxiety-like behaviors similar to a response to chronic restraint stress. These behaviors are concurrent with enhanced synaptic responsiveness and suppressed short- and long-term synaptic plasticity in the longitudinal DG-DG network but not in the transverse DG-CA3 connection. These findings suggest that DG-related anxiety is typified by synaptic alteration in the longitudinal DG-DG network.
Keywords: Behavioral neuroscience; Biological sciences; Neuroscience.
© 2022 The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures





References
-
- Chiba S., Numakawa T., Ninomiya M., Richards M.C., Wakabayashi C., Kunugi H. Chronic restraint stress causes anxiety-and depression-like behaviors, downregulates glucocorticoid receptor expression, and attenuates glutamate release induced by brain-derived neurotrophic factor in the prefrontal cortex. Prog. Neuropsychopharmacol. Biol. Psychiatry. 2012;39:112–119. doi: 10.1016/j.pnpbp.2012.05.018. - DOI - PubMed
LinkOut - more resources
Full Text Sources
Miscellaneous
