

Olfactory Strategies in the Defensive Behaviour of Insects
- PMID: 35621804
- PMCID: PMC9145661
- DOI: 10.3390/insects13050470
Olfactory Strategies in the Defensive Behaviour of Insects
Abstract
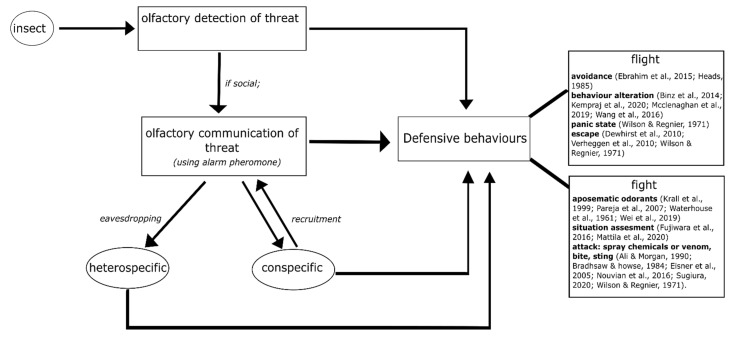
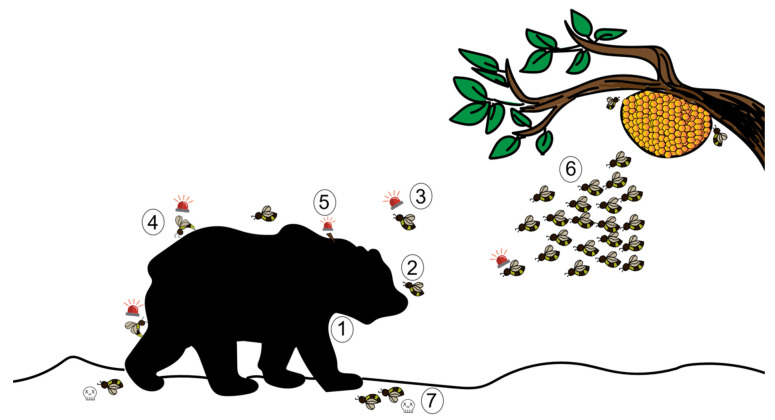
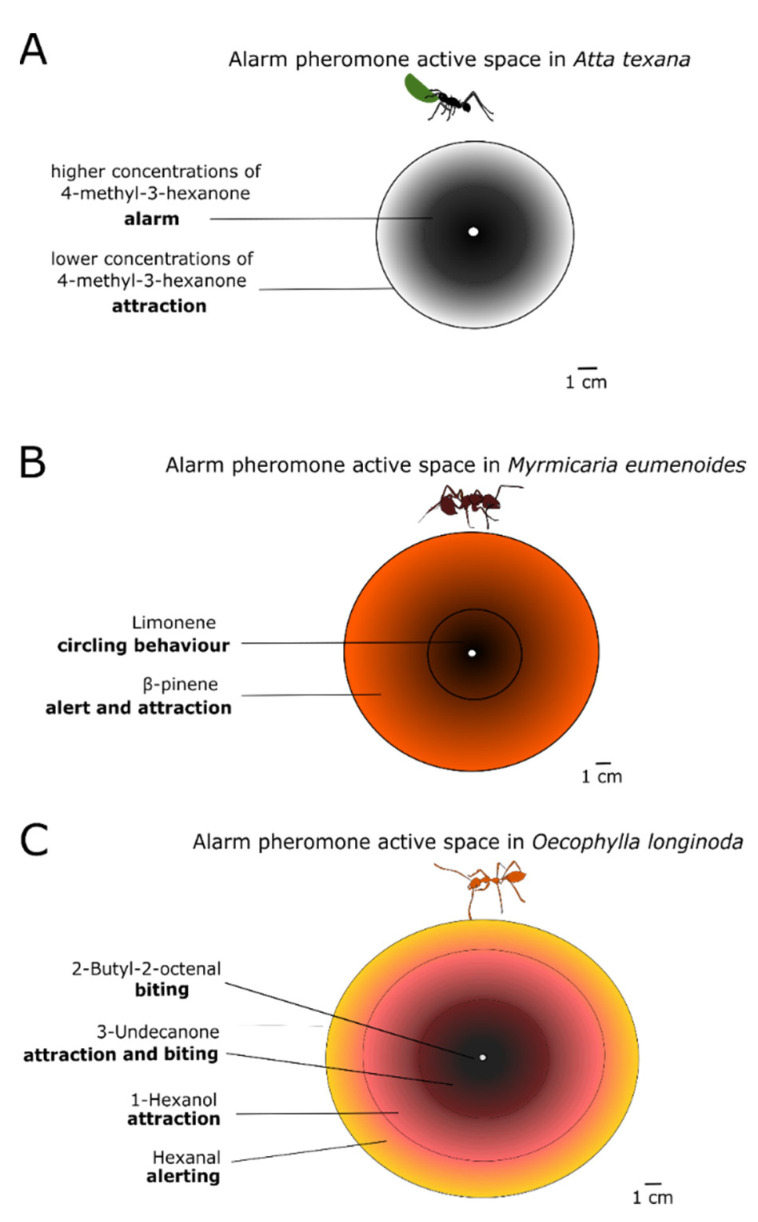
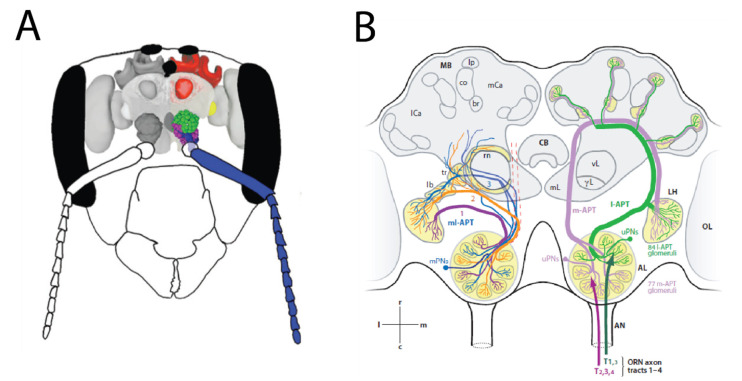
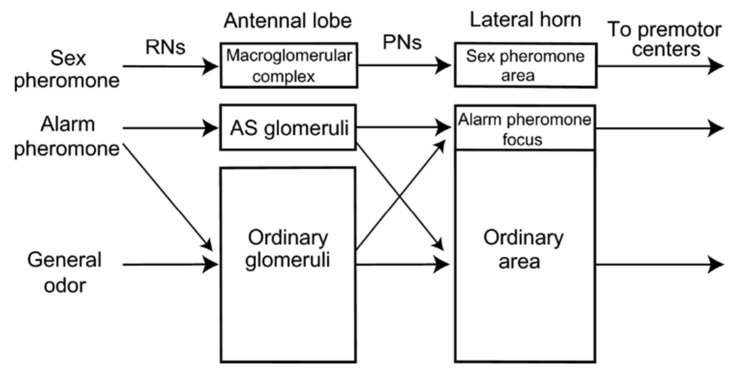
Most animals must defend themselves in order to survive. Defensive behaviour includes detecting predators or intruders, avoiding them by staying low-key or escaping or deterring them away by means of aggressive behaviour, i.e., attacking them. Responses vary across insect species, ranging from individual responses to coordinated group attacks in group-living species. Among different modalities of sensory perception, insects predominantly use the sense of smell to detect predators, intruders, and other threats. Furthermore, social insects, such as honeybees and ants, communicate about danger by means of alarm pheromones. In this review, we focus on how olfaction is put to use by insects in defensive behaviour. We review the knowledge of how chemical signals such as the alarm pheromone are processed in the insect brain. We further discuss future studies for understanding defensive behaviour and the role of olfaction.
Keywords: aggression; alarm pheromone; chemical defense; defensive behaviour; odorant coding; olfactory strategies.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Insect alarm pheromones in response to predators: Ecological trade-offs and molecular mechanisms.Insect Biochem Mol Biol. 2021 Jan;128:103514. doi: 10.1016/j.ibmb.2020.103514. Epub 2020 Dec 23. Insect Biochem Mol Biol. 2021. PMID: 33359575 Review.
-
Honey Bee Alarm Pheromone Mediates Communication in Plant-Pollinator-Predator Interactions.Insects. 2019 Oct 21;10(10):366. doi: 10.3390/insects10100366. Insects. 2019. PMID: 31640201 Free PMC article. Review.
-
Appetitive floral odours prevent aggression in honeybees.Nat Commun. 2015 Dec 22;6:10247. doi: 10.1038/ncomms10247. Nat Commun. 2015. PMID: 26694599 Free PMC article.
-
The Odorant Binding Protein, SiOBP5, Mediates Alarm Pheromone Olfactory Recognition in the Red Imported Fire Ant, Solenopsis invicta.Biomolecules. 2021 Oct 28;11(11):1595. doi: 10.3390/biom11111595. Biomolecules. 2021. PMID: 34827593 Free PMC article.
-
Alarm Pheromone Composition and Behavioral Activity in Fungus-Growing Ants.J Chem Ecol. 2017 Mar;43(3):225-235. doi: 10.1007/s10886-017-0821-4. Epub 2017 Feb 28. J Chem Ecol. 2017. PMID: 28247150 Free PMC article.
Cited by
-
Epidermal oxysterols function as alarm substances in zebrafish.iScience. 2024 Apr 3;27(5):109660. doi: 10.1016/j.isci.2024.109660. eCollection 2024 May 17. iScience. 2024. PMID: 38650983 Free PMC article.
-
Brindley's Glands Volatilome of the Predator Zelus renardii Interacting with Xylella Vectors.Insects. 2023 Jun 3;14(6):520. doi: 10.3390/insects14060520. Insects. 2023. PMID: 37367336 Free PMC article.
-
iORandLigandDB: A Website for Three-Dimensional Structure Prediction of Insect Odorant Receptors and Docking with Odorants.Insects. 2023 Jun 15;14(6):560. doi: 10.3390/insects14060560. Insects. 2023. PMID: 37367376 Free PMC article.
-
Microbial metabolites as engines of behavioral variation across animals.Gut Microbes. 2025 Dec;17(1):2501191. doi: 10.1080/19490976.2025.2501191. Epub 2025 May 13. Gut Microbes. 2025. PMID: 40357979 Free PMC article. Review.
-
Autofluorescent Biomolecules in Diptera: From Structure to Metabolism and Behavior.Molecules. 2022 Jul 12;27(14):4458. doi: 10.3390/molecules27144458. Molecules. 2022. PMID: 35889334 Free PMC article. Review.
References
-
- Lima S.L., Dill L.M. Behavioral decisions made under the risk of predation: A review and prospectus. Can. J. Zool. 1990;68:619–640. doi: 10.1139/z90-092. - DOI
-
- Heads P.A. The effect of invertebrate and vertebrate predators on the foraging movements of Ischnura elegans larvae (Odonata: Zygoptera) Freshw. Biol. 1985;15:559–571. doi: 10.1111/j.1365-2427.1985.tb00226.x. - DOI
-
- Binz H., Bucher R., Entling M.H., Menzel F. Knowing the Risk: Crickets Distinguish between Spider Predators of Different Size and Commonness. Ethology. 2014;120:99–110. doi: 10.1111/eth.12183. - DOI
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
