

Dysregulation and therapeutic targeting of RNA splicing in cancer
- PMID: 35624337
- PMCID: PMC9551392
- DOI: 10.1038/s43018-022-00384-z
Dysregulation and therapeutic targeting of RNA splicing in cancer
Abstract
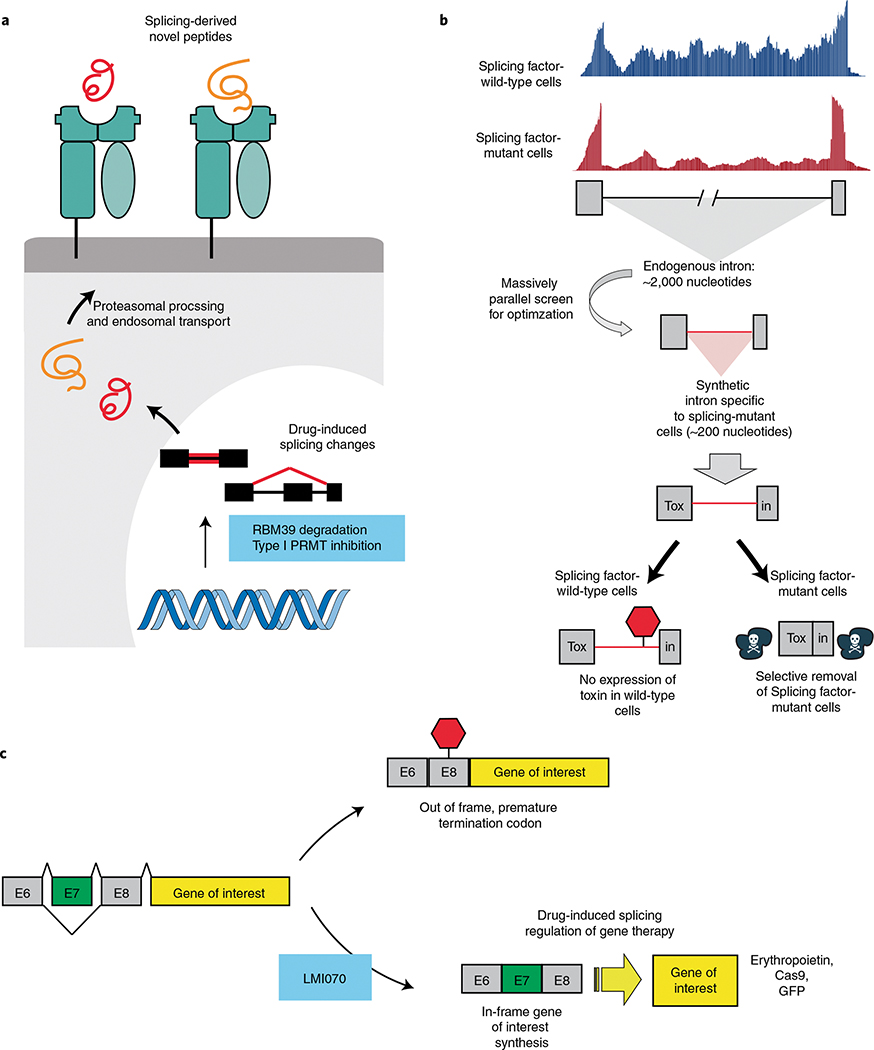
High-throughput sequencing and functional characterization of the cancer transcriptome have uncovered cancer-specific dysregulation of RNA splicing across a variety of cancers. Alterations in the cancer genome and dysregulation of RNA splicing factors lead to missplicing, splicing alteration-dependent gene expression and, in some cases, generation of novel splicing-derived proteins. Here, we review recent advances in our understanding of aberrant splicing in cancer pathogenesis and present strategies to harness cancer-specific aberrant splicing for therapeutic intent.
© 2022. Springer Nature America, Inc.
Conflict of interest statement
Competing interests
O.A.-W. has served as a consultant for H3B Biomedicine, Foundation Medicine, Inc., Merck, Prelude Therapeutics and Janssen and is on the Scientific Advisory Board of Envisagenics, Inc., AIChemy, Harmonic Discovery, Inc., and Pfizer Boulder. O.A.-W. has received prior research funding from H3B Biomedicine, Nurix Therapeutics and LOXO Oncology unrelated to the current manuscript. The remaining authors declare no competing interests.
Figures



References
-
- Brody E & Abelson J The ‘spliceosome’: yeast pre-messenger RNA associates with a 40S complex in a splicing-dependent reaction. Science 228, 963–967 (1985). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
