

Antagonizing Glutamine Bioavailability Promotes Radiation Sensitivity in Prostate Cancer
- PMID: 35626095
- PMCID: PMC9139225
- DOI: 10.3390/cancers14102491
Antagonizing Glutamine Bioavailability Promotes Radiation Sensitivity in Prostate Cancer
Abstract
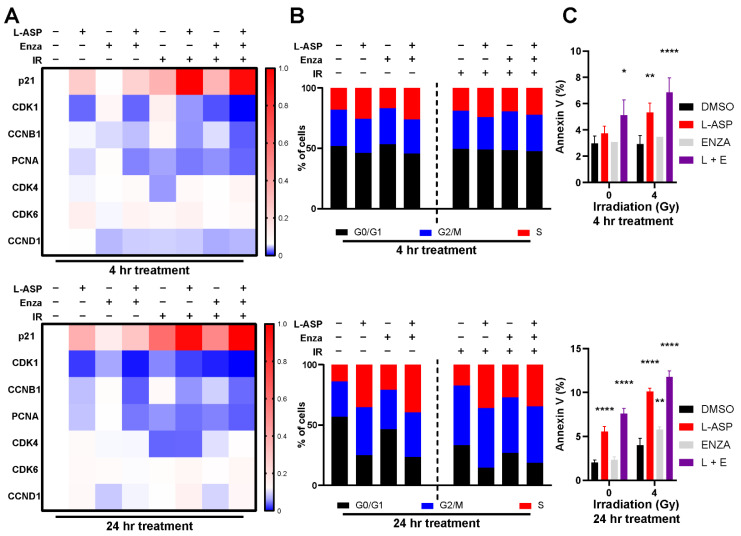
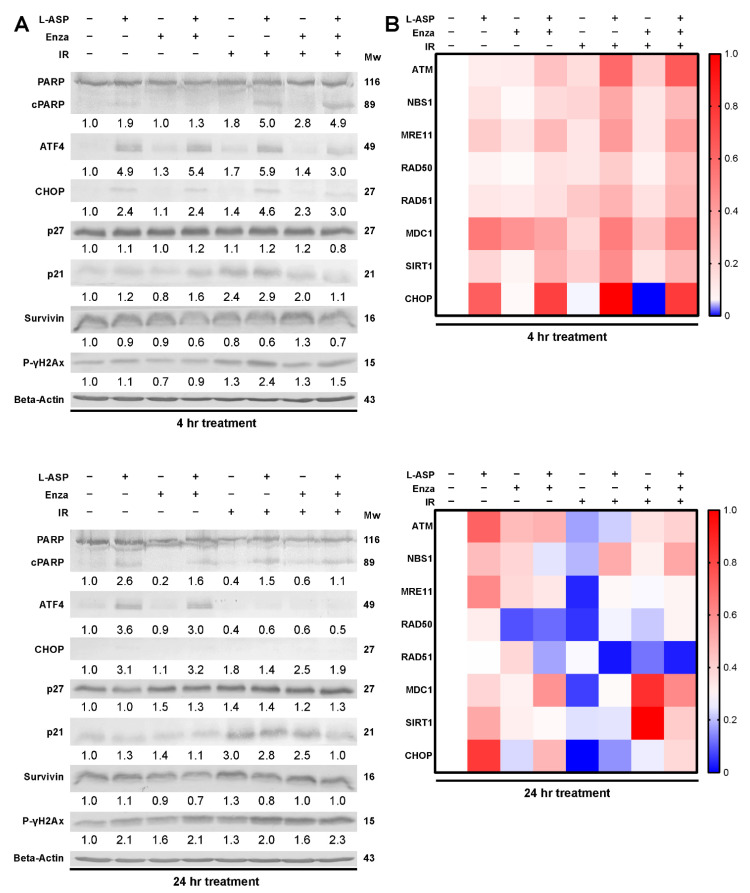
Nearly half of localized prostate cancer (PCa) patients given radiation therapy develop recurrence. Here, we identified glutamine as a key player in mediating the radio-sensitivity of PCa. Glutamine transporters and glutaminase are upregulated by radiation therapy of PCa cells, but respective inhibitors were ineffective in radio-sensitization. However, targeting glutamine bioavailability by L-asparaginase (L-ASP) led to a significant reduction in clonogenicity when combined with irradiation. L-ASP reduced extracellular asparagine and glutamine, but the sensitization effects were driven through its depletion of glutamine. L-ASP led to G2/M cell cycle checkpoint blockade. As evidence, there was a respective delay in DNA repair associated with RAD51 downregulation and upregulation of CHOP, contributing to radiation-induced cell death. A radio-resistant PCa cell line was developed, was found to bypass radiation-induced mitotic catastrophe, and was sensitive to L-ASP/radiation combination treatment. Previously, PCa-associated fibroblasts were reported as a glutamine source supporting tumor progression. As such, glutamine-free media were not effective in promoting radiation-induced PCa cell death when co-cultured with associated primary fibroblasts. However, the administration L-ASP catalyzed glutamine depletion with irradiated co-cultures and catalyzed tumor volume reduction in a mouse model. The clinical history of L-ASP for leukemia patients supports the viability for its repurposing as a radio-sensitizer for PCa patients.
Keywords: asparaginase; glutamine; prostate cancer; radiation therapy.
Conflict of interest statement
The authors declare no conflict of interest.
Figures




Similar articles
-
Antagonizing CD105 enhances radiation sensitivity in prostate cancer.Oncogene. 2018 Aug;37(32):4385-4397. doi: 10.1038/s41388-018-0278-0. Epub 2018 May 2. Oncogene. 2018. PMID: 29717261 Free PMC article.
-
L-asparaginase-mediated Therapy in L-asparagine Auxotrophic Cancers: A Review.Anticancer Agents Med Chem. 2022;22(13):2393-2410. doi: 10.2174/1871520622666220106103336. Anticancer Agents Med Chem. 2022. PMID: 34994334 Review.
-
Correlation of L-asp Activity, Anti-L-asp Antibody, Asn and Gln With Adverse Events Especially Anaphylaxis Risks in PEG-asp-Contained Regime Treated Pediatric ALL Patients.Technol Cancer Res Treat. 2020 Jan-Dec;19:1533033820980113. doi: 10.1177/1533033820980113. Technol Cancer Res Treat. 2020. PMID: 33287663 Free PMC article.
-
Glutaminolysis is a metabolic route essential for survival and growth of prostate cancer cells and a target of 5α-dihydrotestosterone regulation.Cell Oncol (Dordr). 2021 Apr;44(2):385-403. doi: 10.1007/s13402-020-00575-9. Epub 2021 Jan 19. Cell Oncol (Dordr). 2021. PMID: 33464483
-
ω-Amidase: an underappreciated, but important enzyme in L-glutamine and L-asparagine metabolism; relevance to sulfur and nitrogen metabolism, tumor biology and hyperammonemic diseases.Amino Acids. 2016 Jan;48(1):1-20. doi: 10.1007/s00726-015-2061-7. Epub 2015 Aug 11. Amino Acids. 2016. PMID: 26259930 Review.
Cited by
-
Glutamine Metabolism and Prostate Cancer.Cancers (Basel). 2024 Aug 18;16(16):2871. doi: 10.3390/cancers16162871. Cancers (Basel). 2024. PMID: 39199642 Free PMC article. Review.
-
Stromal-Epithelial Interactions in Cancer Progression and Therapy Response.Cancers (Basel). 2023 Jun 1;15(11):3014. doi: 10.3390/cancers15113014. Cancers (Basel). 2023. PMID: 37296976 Free PMC article.
-
A Five Glutamine-Associated Signature Predicts Prognosis of Prostate Cancer and Links Glutamine Metabolism with Tumor Microenvironment.J Clin Med. 2023 Mar 14;12(6):2243. doi: 10.3390/jcm12062243. J Clin Med. 2023. PMID: 36983244 Free PMC article.
-
Identification of prognostic hub genes and functional role of BAIAP2L2 in prostate cancer progression: a transcriptomic and experimental study.Front Immunol. 2025 Apr 28;16:1543476. doi: 10.3389/fimmu.2025.1543476. eCollection 2025. Front Immunol. 2025. PMID: 40356901 Free PMC article.
-
Cellular Signaling of Amino Acid Metabolism in Prostate Cancer.Int J Mol Sci. 2025 Jan 17;26(2):776. doi: 10.3390/ijms26020776. Int J Mol Sci. 2025. PMID: 39859489 Free PMC article. Review.
References
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
