

Understanding the Dynamics of the Transient and Permanent Opening Events of the Mitochondrial Permeability Transition Pore with a Novel Stochastic Model
- PMID: 35629820
- PMCID: PMC9146742
- DOI: 10.3390/membranes12050494
Understanding the Dynamics of the Transient and Permanent Opening Events of the Mitochondrial Permeability Transition Pore with a Novel Stochastic Model
Abstract
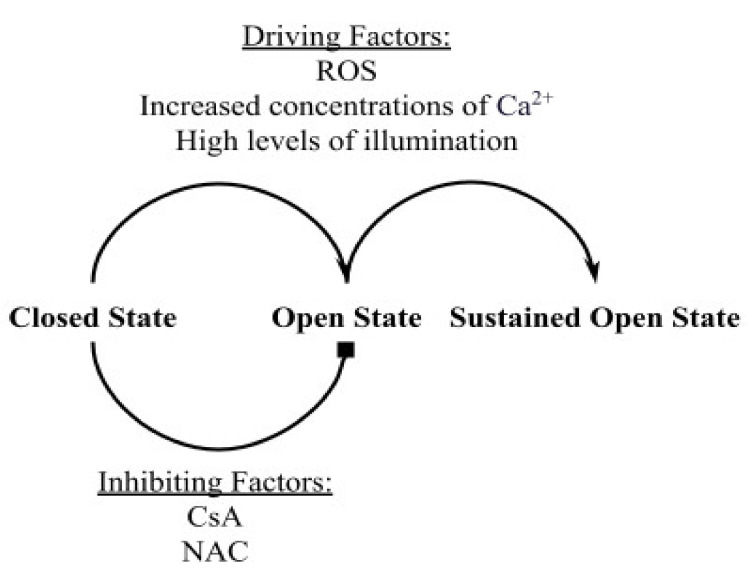
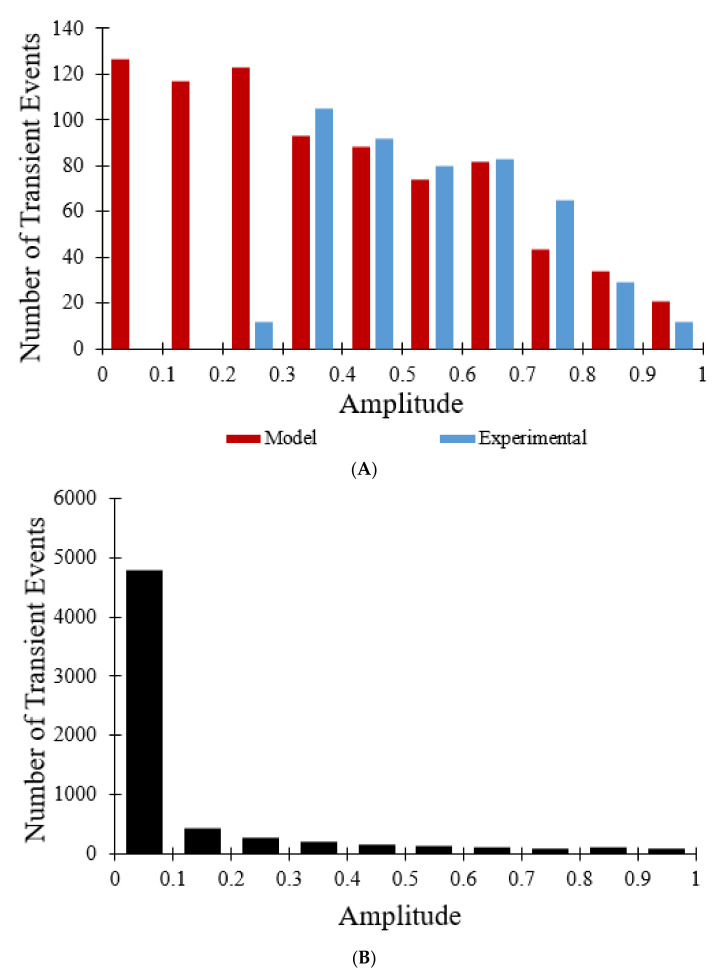
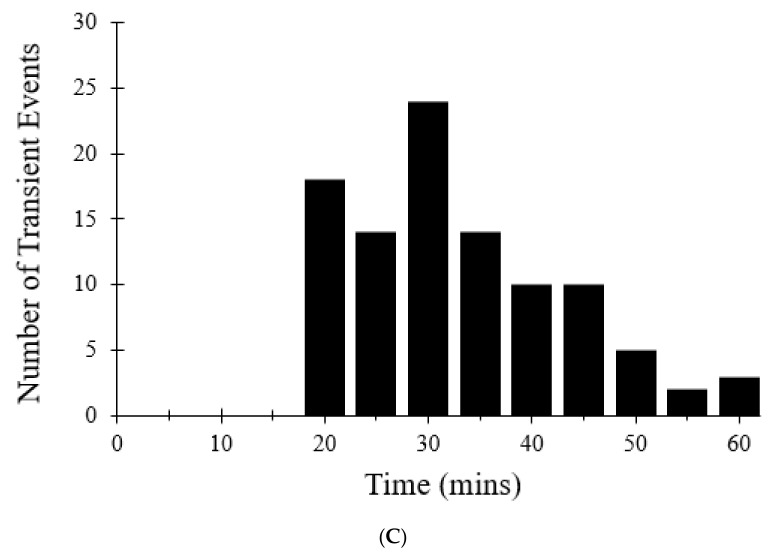
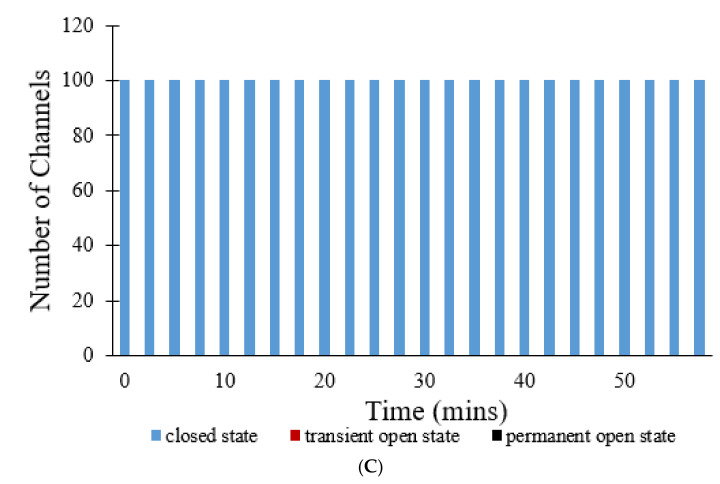
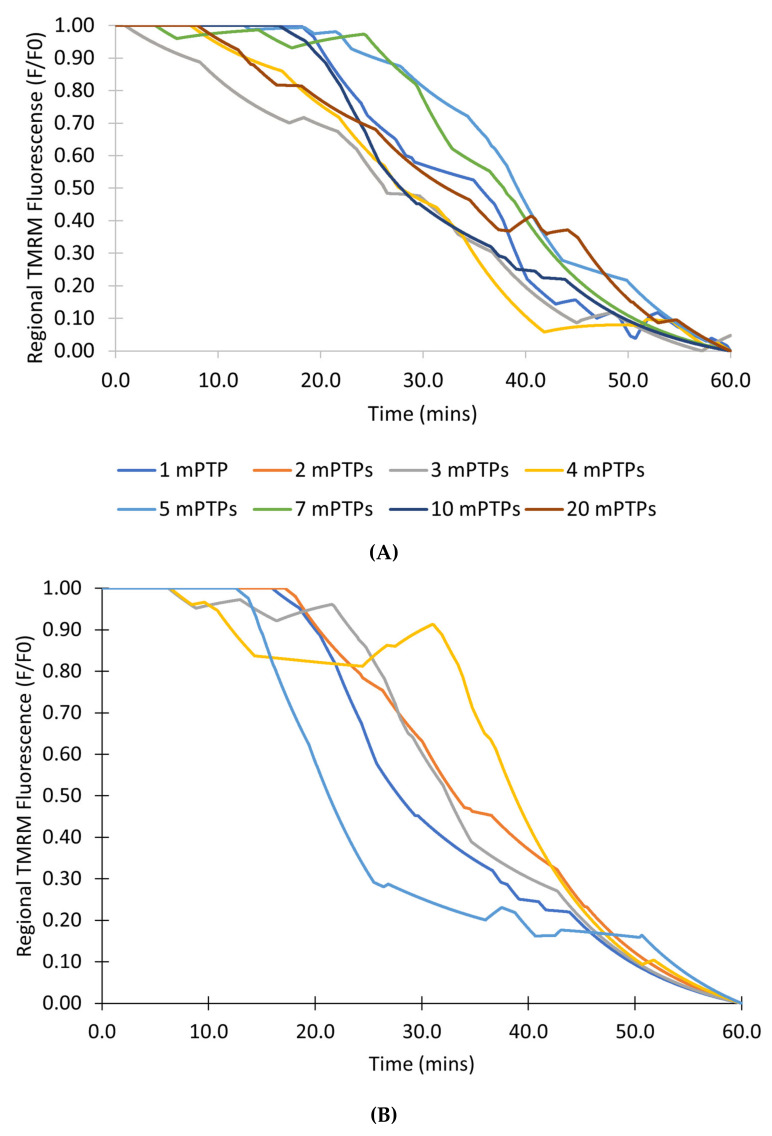
The mitochondrial permeability transition pore (mPTP) is a non-selective pore in the inner mitochondrial membrane (IMM) which causes depolarization when it opens under conditions of oxidative stress and high concentrations of Ca2+. In this study, a stochastic computational model was developed to better understand the dynamics of mPTP opening and closing associated with elevated reactive oxygen species (ROS) in cardiomyocytes. The data modeled are from "photon stress" experiments in which the fluorescent dye TMRM (tetramethylrhodamine methyl ester) is both the source of ROS (induced by laser light) and sensor of the electrical potential difference across the IMM. Monte Carlo methods were applied to describe opening and closing of the pore along with the Hill Equation to account for the effect of ROS levels on the transition probabilities. The amplitude distribution of transient mPTP opening events, the number of transient mPTP opening events per minute in a cell, the time it takes for recovery after transient depolarizations in the mitochondria, and the change in TMRM fluorescence during the transition from transient to permanent mPTP opening events were analyzed. The model suggests that mPTP transient open times have an exponential distribution that are reflected in TMRM fluorescence. A second multiple pore model in which individual channels have no permanent open state suggests that 5-10 mPTP per mitochondria would be needed for sustained mitochondrial depolarization at elevated ROS with at least 1 mPTP in the transient open state.
Keywords: Monte Carlo method; TMRM; computational model; depolarization; heart; mPTP; mitochondria; reactive oxygen species (ROS).
Conflict of interest statement
The authors declare no conflict of interest.
Figures











References
-
- Luis-García E.R., Becerril C., Salgado-Aguayo A., Aparicio-Trejo O.E., Romero Y., Flores-Soto E., Mendoza-Milla C., Montaño M., Chagoya V., Pedraza-Chaverri J., et al. Mitochondrial Dysfunction and Alterations in Mitochondrial Permeability Transition Pore (mPTP) Contribute to Apoptosis Resistance in Idiopathic Pulmonary Fibrosis Fibroblasts. Int. J. Mol. Sci. 2021;22:7870. doi: 10.3390/ijms22157870. - DOI - PMC - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
