

BCL6-dependent TCF-1+ progenitor cells maintain effector and helper CD4+ T cell responses to persistent antigen
- PMID: 35637103
- PMCID: PMC10034764
- DOI: 10.1016/j.immuni.2022.05.003
BCL6-dependent TCF-1+ progenitor cells maintain effector and helper CD4+ T cell responses to persistent antigen
Abstract
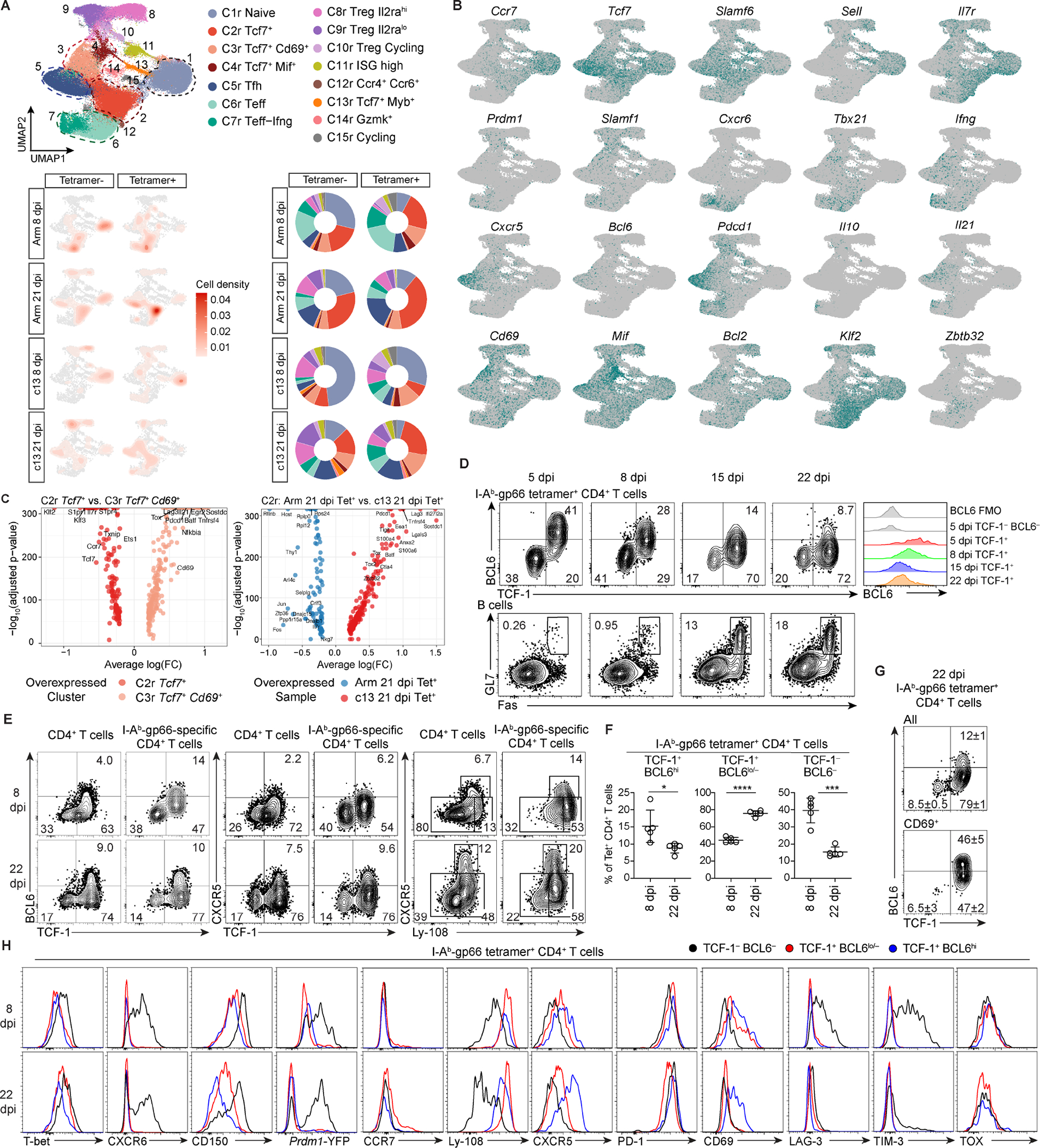
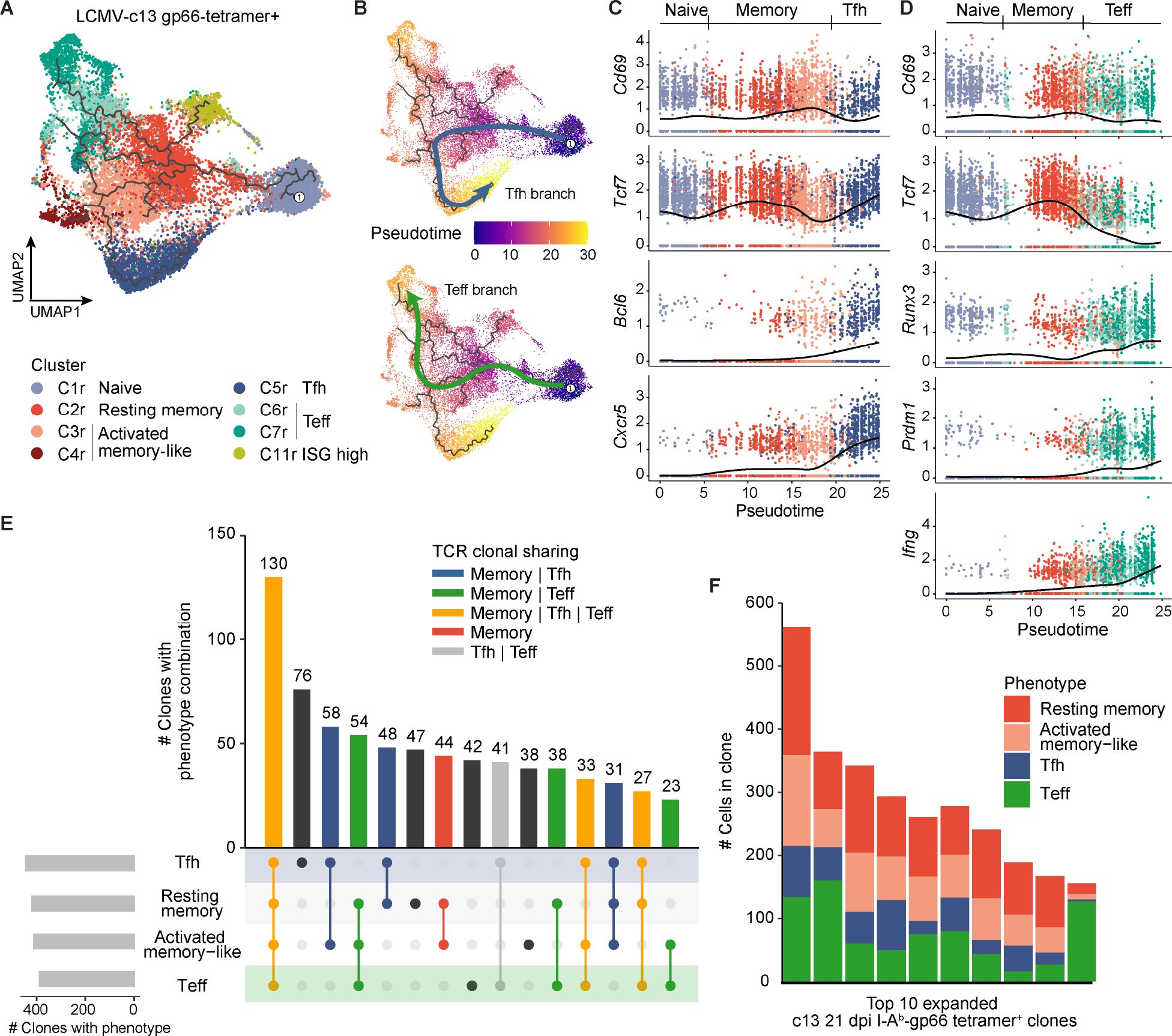
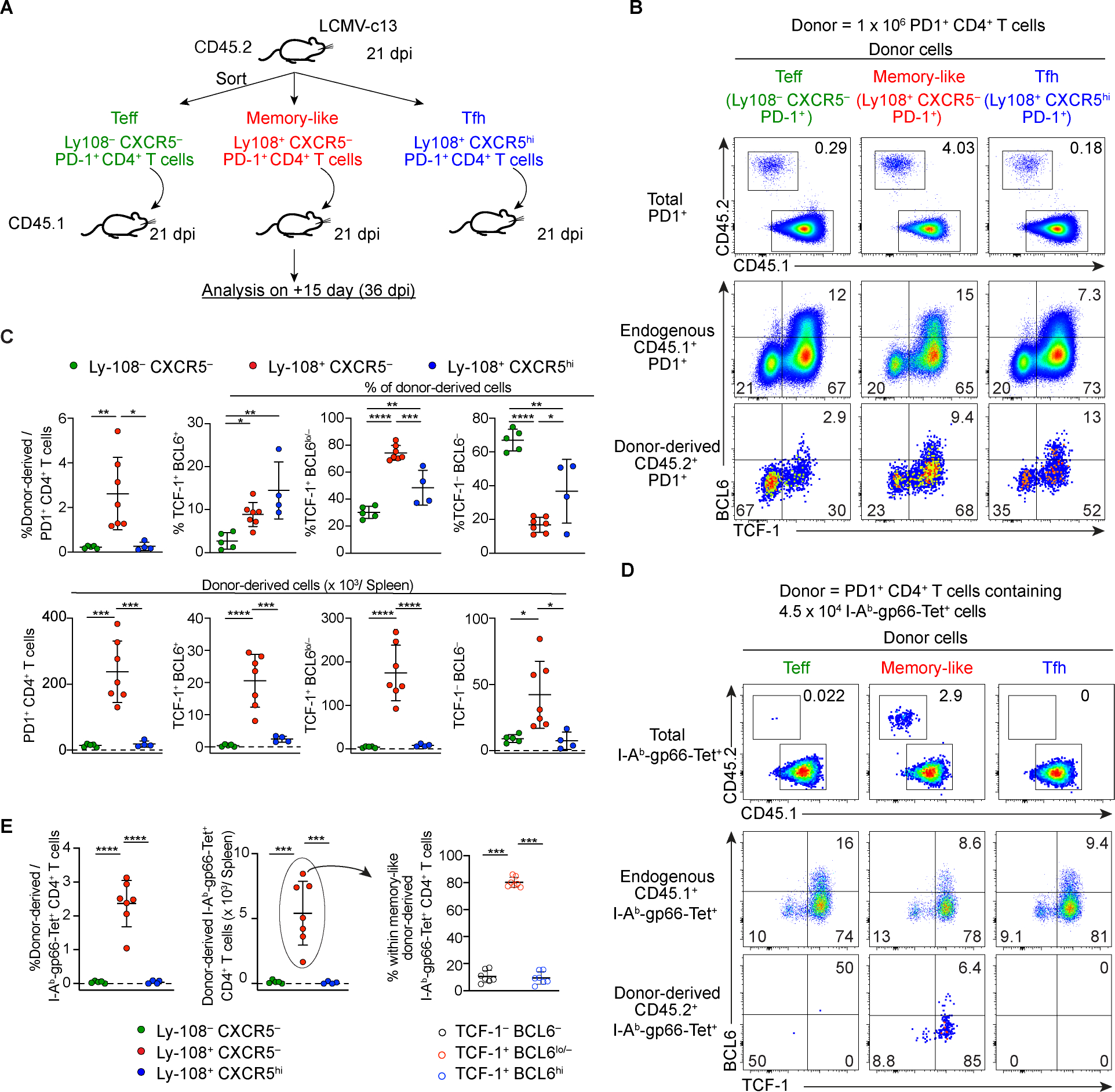
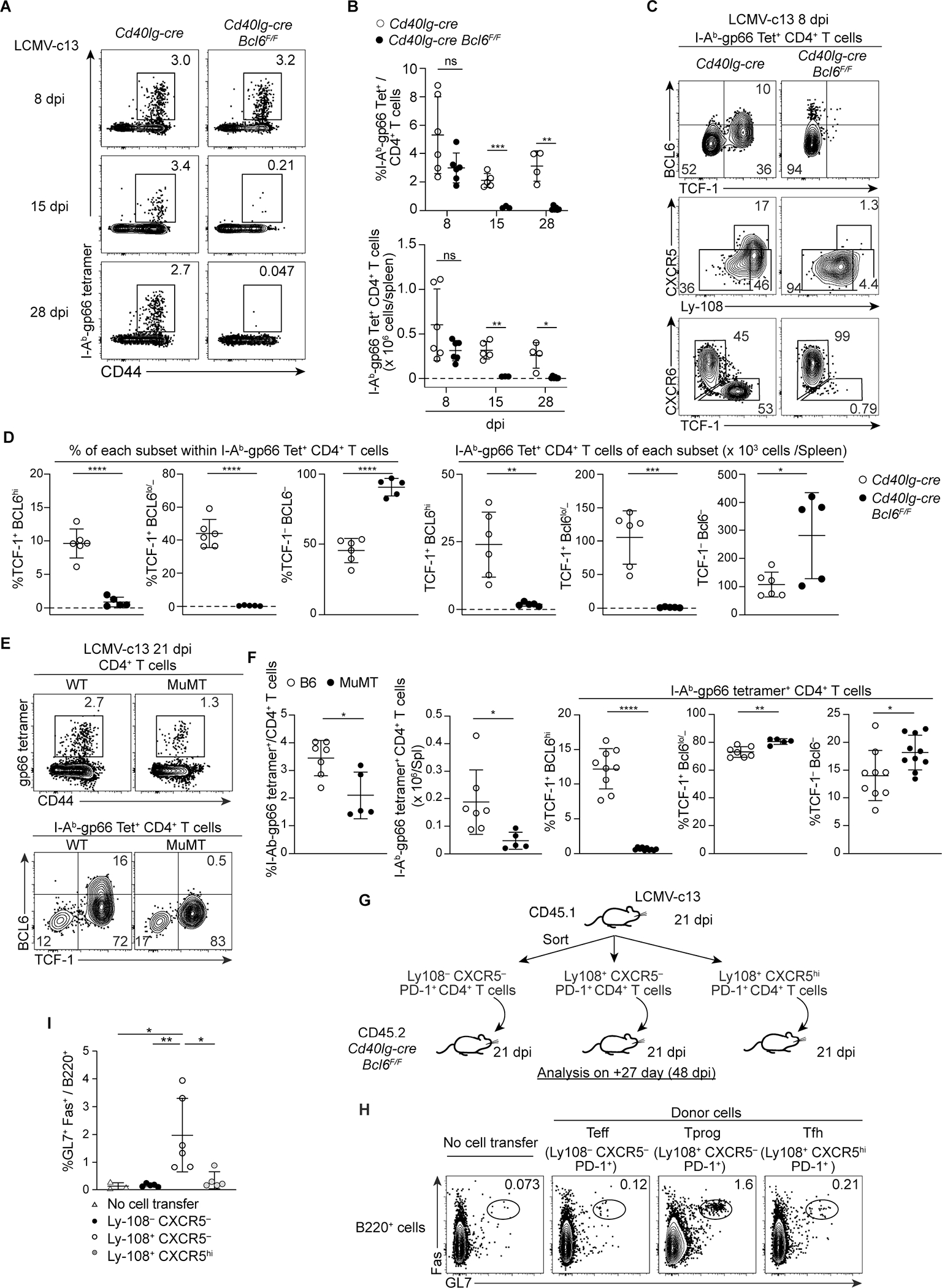
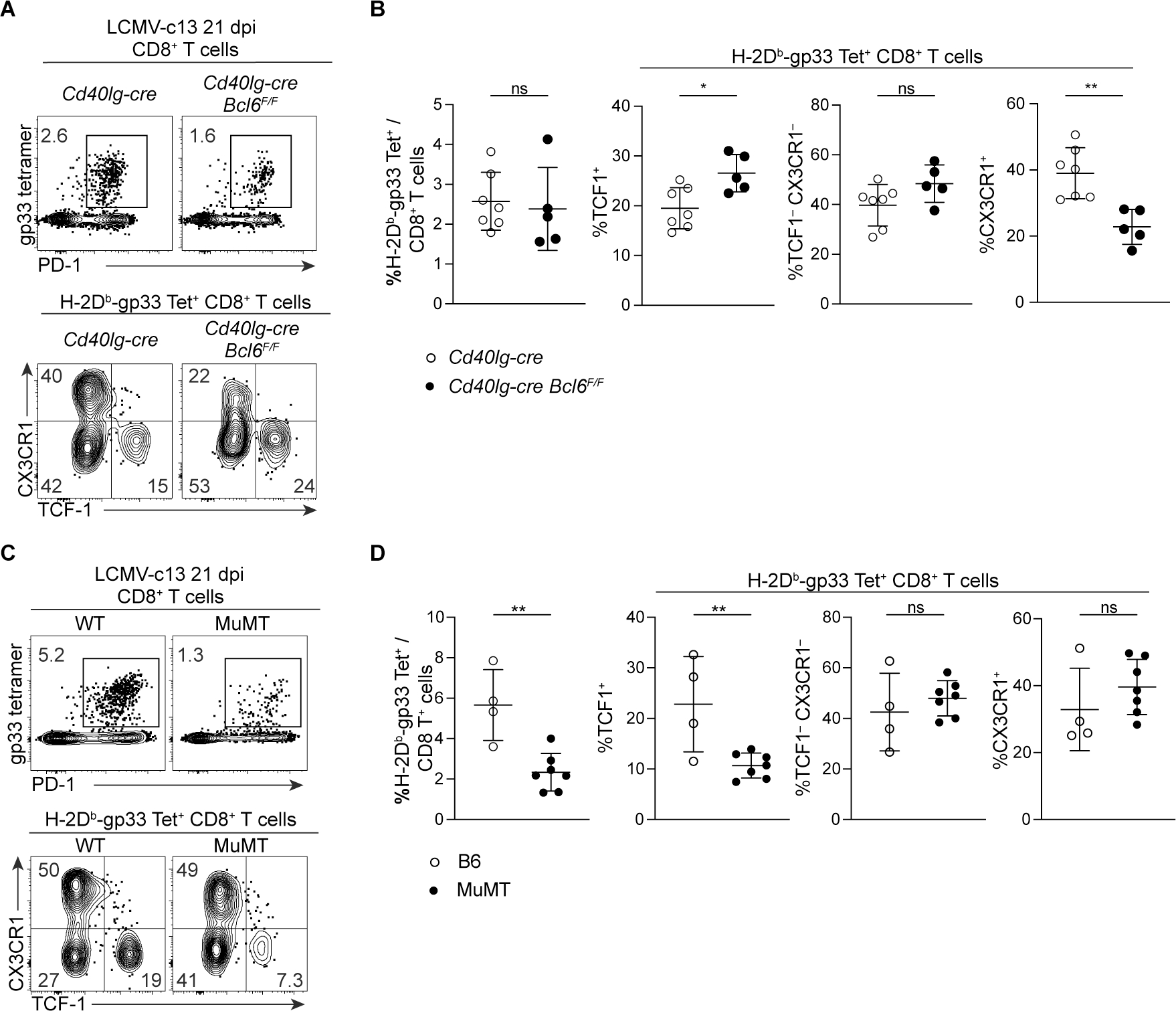
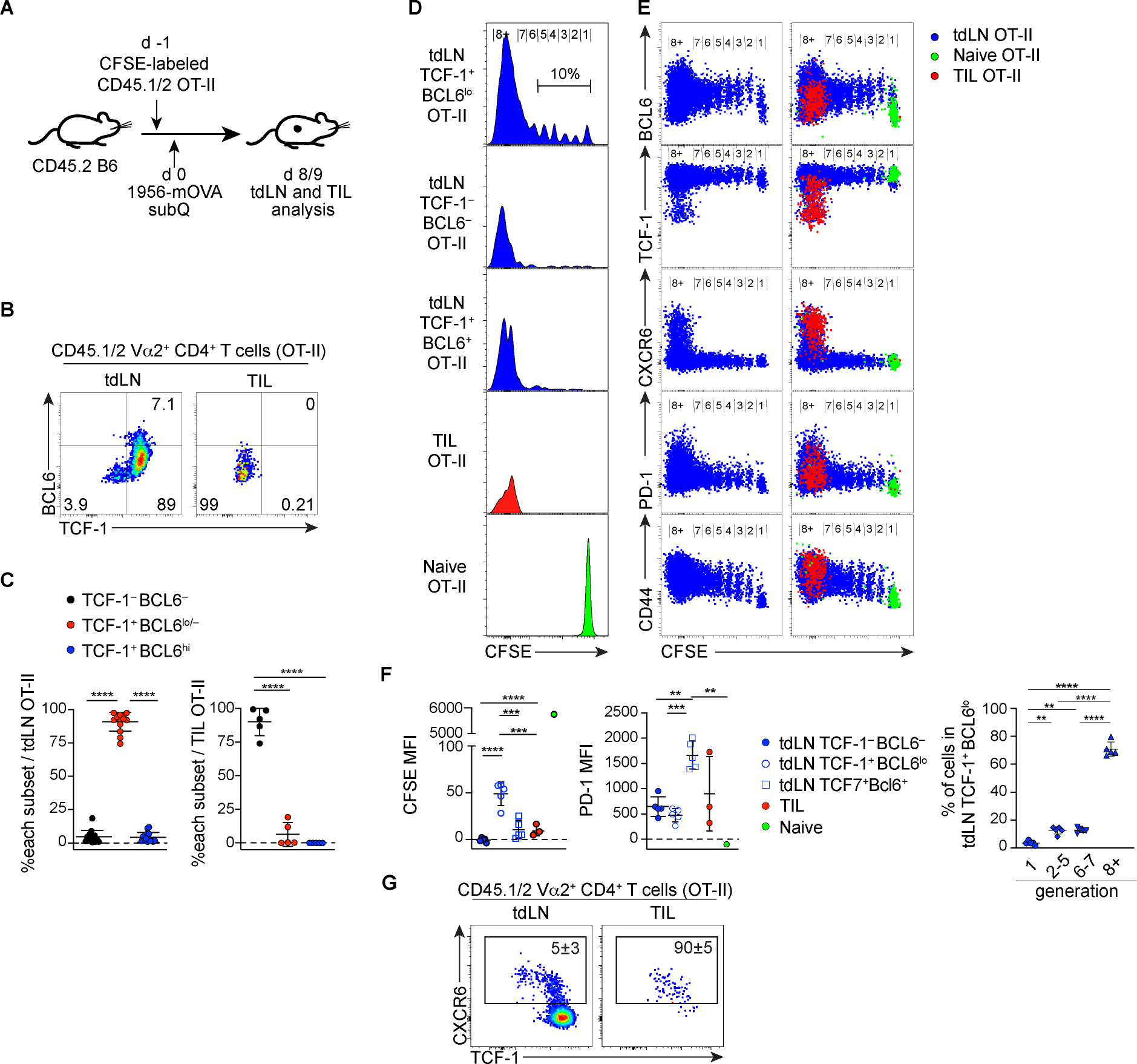
Soon after activation, CD4+ T cells are segregated into BCL6+ follicular helper (Tfh) and BCL6- effector (Teff) T cells. Here, we explored how these subsets are maintained during chronic antigen stimulation using the mouse chronic LCMV infection model. Using single cell-transcriptomic and epigenomic analyses, we identified a population of PD-1+ TCF-1+ CD4+ T cells with memory-like features. TCR clonal tracing and adoptive transfer experiments demonstrated that these cells have self-renewal capacity and continue to give rise to both Teff and Tfh cells, thus functioning as progenitor cells. Conditional deletion experiments showed Bcl6-dependent development of these progenitors, which were essential for sustaining antigen-specific CD4+ T cell responses to chronic infection. An analogous CD4+ T cell population developed in draining lymph nodes in response to tumors. Our study reveals the heterogeneity and plasticity of CD4+ T cells during persistent antigen exposure and highlights their population dynamics through a stable, bipotent intermediate state.
Keywords: CD4(+) T cells; anti-tumor immunity; chronic viral infection; effector T cells; follicular helper T cells; progenitor cells; single-cell genomics.
Copyright © 2022 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests A.T.S. is a scientific co-founder of Immunai, founder of Cartography Biosciences, and receives research funding from Arsenal Biosciences, Merck Research Laboratories, and Allogene Therapeutics.
Figures

Comment in
-
CD4+ progenitor cells sustain helper responses during chronic infection.Immunity. 2022 Jul 12;55(7):1143-1145. doi: 10.1016/j.immuni.2022.06.016. Immunity. 2022. PMID: 35830820
References
-
- Baumjohann D, Okada T, and Ansel KM (2011). Cutting Edge: Distinct waves of BCL6 expression during T follicular helper cell development. J. Immunol. 187, 2089–2092. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
