

Switches, stability and reversals in the evolutionary history of sexual systems in fish
- PMID: 35637181
- PMCID: PMC9151764
- DOI: 10.1038/s41467-022-30419-z
Switches, stability and reversals in the evolutionary history of sexual systems in fish
Abstract
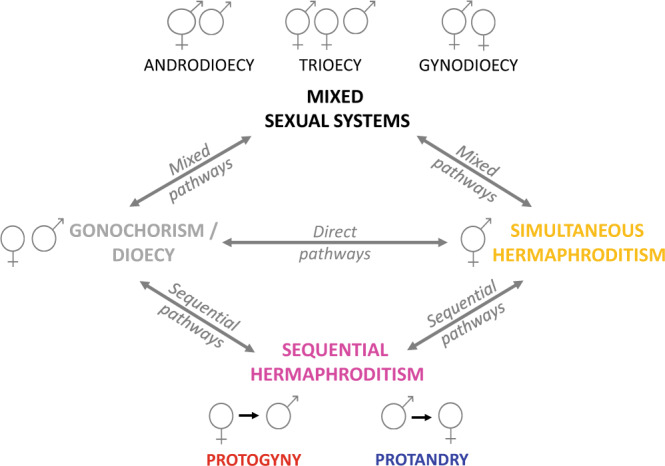
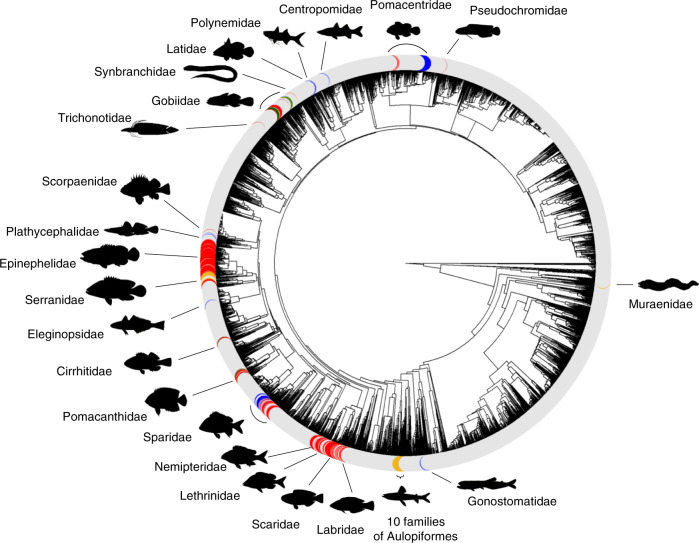
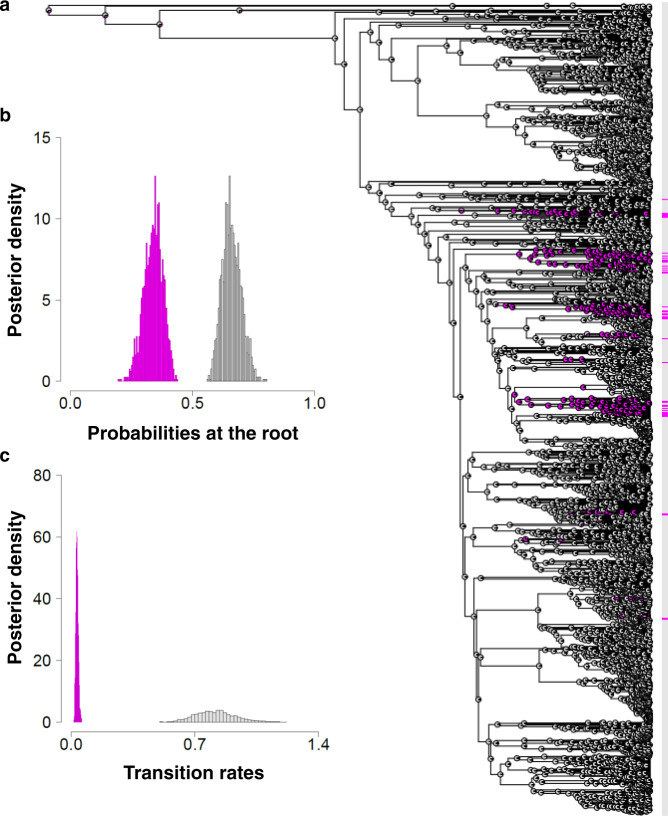
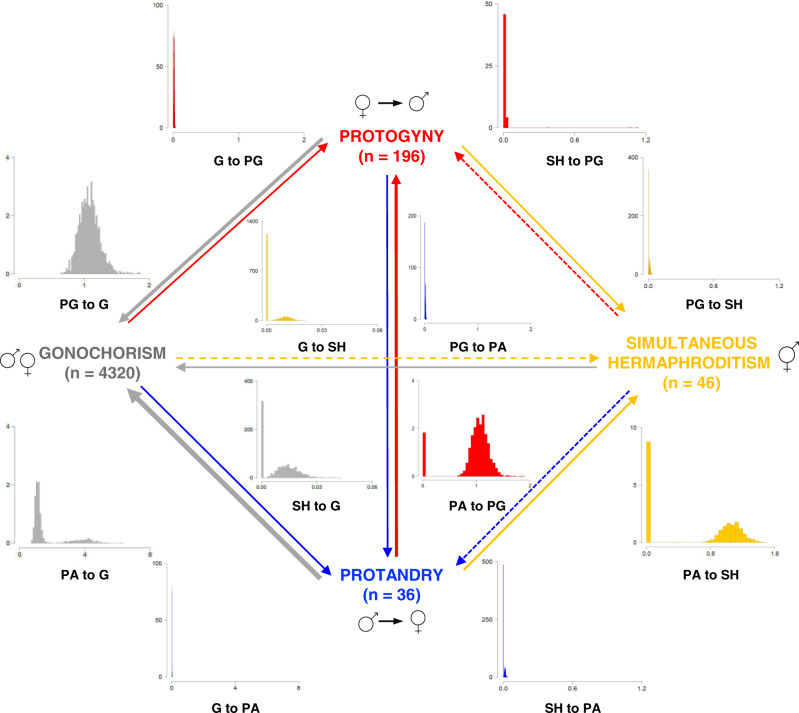
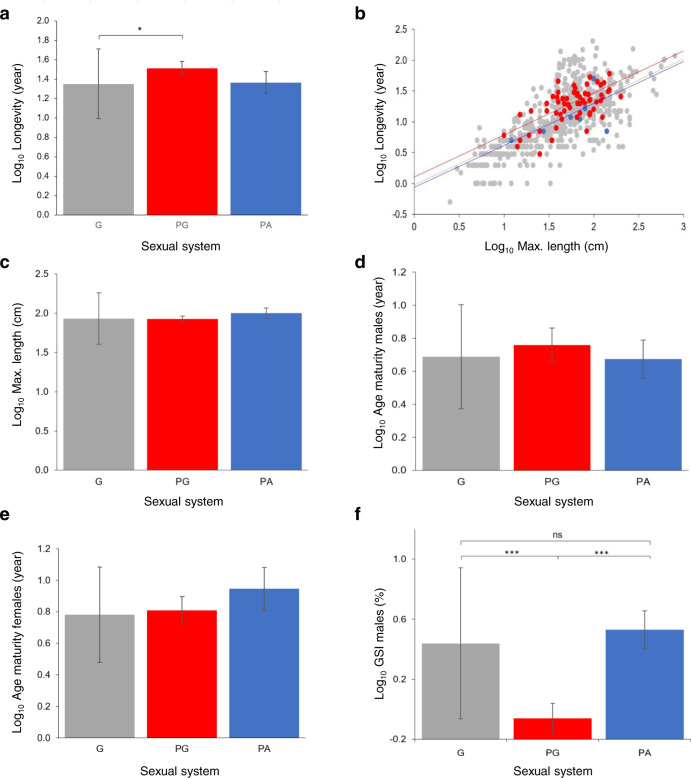
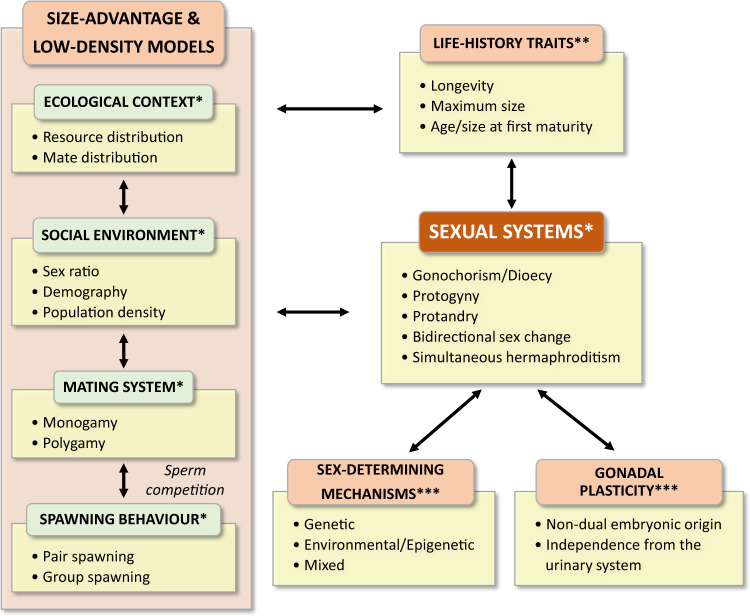
Sexual systems are highly diverse and have profound consequences for population dynamics and resilience. Yet, little is known about how they evolved. Using phylogenetic Bayesian modelling and a sample of 4614 species, we show that gonochorism is the likely ancestral condition in teleost fish. While all hermaphroditic forms revert quickly to gonochorism, protogyny and simultaneous hermaphroditism are evolutionarily more stable than protandry. In line with theoretical expectations, simultaneous hermaphroditism does not evolve directly from gonochorism but can evolve slowly from sequential hermaphroditism, particularly protandry. We find support for the predictions from life history theory that protogynous, but not protandrous, species live longer than gonochoristic species and invest the least in male gonad mass. The distribution of teleosts' sexual systems on the tree of life does not seem to reflect just adaptive predictions, suggesting that adaptations alone may not fully explain why some sexual forms evolve in some taxa but not others (Williams' paradox). We propose that future studies should incorporate mating systems, spawning behaviours, and the diversity of sex determining mechanisms. Some of the latter might constrain the evolution of hermaphroditism, while the non-duality of the embryological origin of teleost gonads might explain why protogyny predominates over protandry in teleosts.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Leonard, J. L. The evolution of sexual systems in animals. In: Leonard, J.L. (ed.). Transitions between sexual systems: understanding the mechanisms of, and pathways between, dioecy, hermaphroditism and other sexual systems, 1–58 Springer (2019).
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
