

InvL, an Invasin-Like Adhesin, Is a Type II Secretion System Substrate Required for Acinetobacter baumannii Uropathogenesis
- PMID: 35638734
- PMCID: PMC9245377
- DOI: 10.1128/mbio.00258-22
InvL, an Invasin-Like Adhesin, Is a Type II Secretion System Substrate Required for Acinetobacter baumannii Uropathogenesis
Abstract
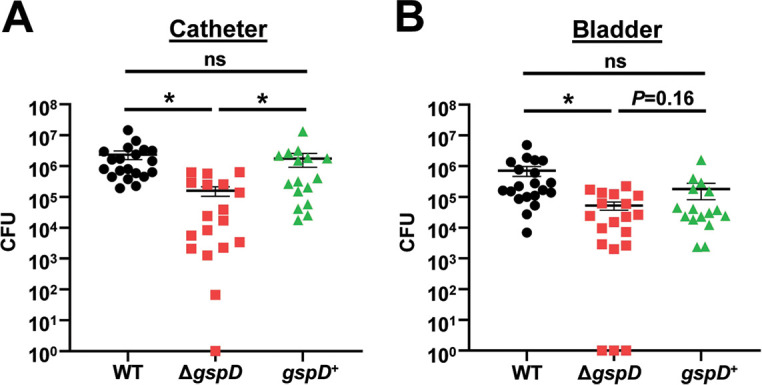
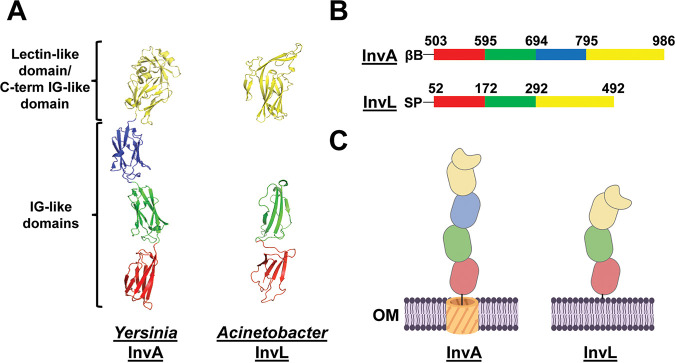
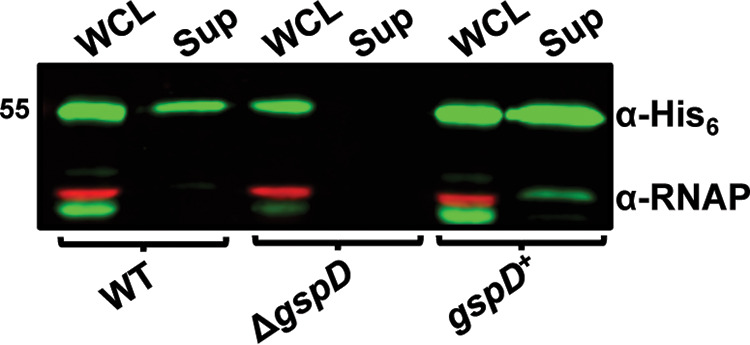
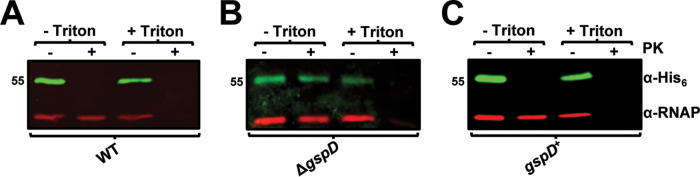
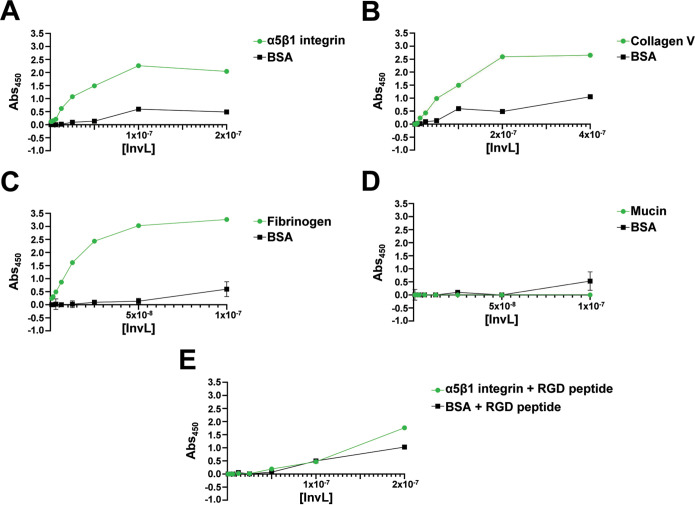
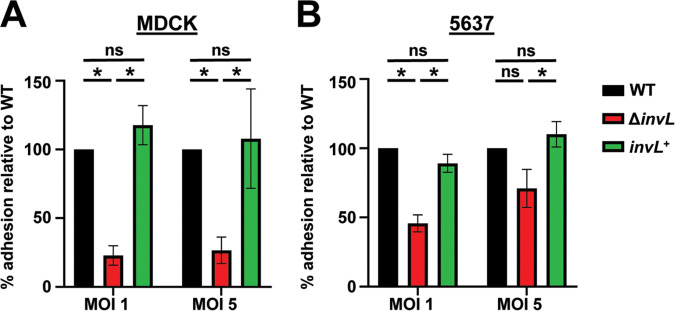
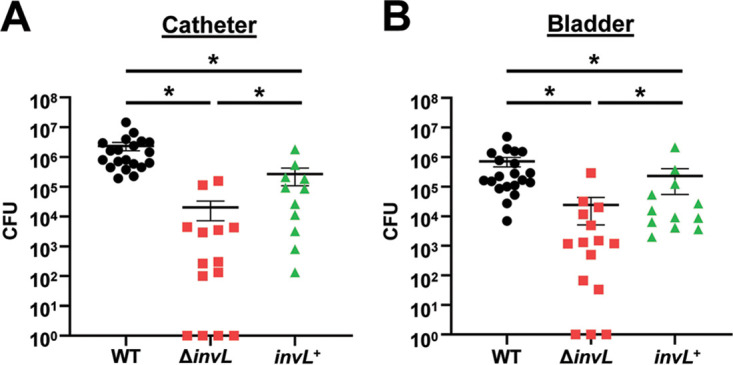
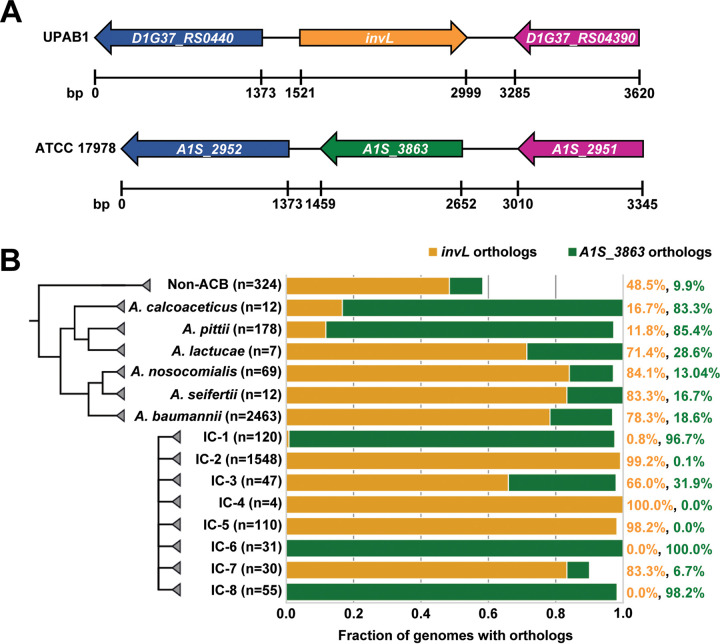
Acinetobacter baumannii is an opportunistic pathogen of growing concern, as isolates are commonly multidrug resistant. While A. baumannii is most frequently associated with pulmonary infections, a significant proportion of clinical isolates come from urinary sources, highlighting its uropathogenic potential. The type II secretion system (T2SS) of commonly used model Acinetobacter strains is important for virulence in various animal models, but the potential role of the T2SS in urinary tract infection (UTI) remains unknown. Here, we used a catheter-associated UTI (CAUTI) model to demonstrate that a modern urinary isolate, UPAB1, requires the T2SS for full virulence. A proteomic screen to identify putative UPAB1 T2SS effectors revealed an uncharacterized lipoprotein with structural similarity to the intimin-invasin family, which serve as type V secretion system (T5SS) adhesins required for the pathogenesis of several bacteria. This protein, designated InvL, lacked the β-barrel domain associated with T5SSs but was confirmed to require the T2SS for both surface localization and secretion. This makes InvL the first identified T2SS effector belonging to the intimin-invasin family. InvL was confirmed to be an adhesin, as the protein bound to extracellular matrix components and mediated adhesion to urinary tract cell lines in vitro. Additionally, the invL mutant was attenuated in the CAUTI model, indicating a role in Acinetobacter uropathogenesis. Finally, bioinformatic analyses revealed that InvL is present in nearly all clinical isolates belonging to international clone 2, a lineage of significant clinical importance. In all, we conclude that the T2SS substrate InvL is an adhesin required for A. baumannii uropathogenesis. IMPORTANCE While pathogenic Acinetobacter can cause various infections, we recently found that 20% of clinical isolates come from urinary sources. Despite the clinical relevance of Acinetobacter as a uropathogen, few virulence factors involved in urinary tract colonization have been defined. Here, we identify a novel type II secretion system effector, InvL, which is required for full uropathogenesis by a modern urinary isolate. Although InvL has predicted structural similarity to the intimin-invasin family of autotransporter adhesins, InvL is predicted to be anchored to the membrane as a lipoprotein. Similar to other invasin homologs, however, we demonstrate that InvL is a bona fide adhesin capable of binding extracellular matrix components and mediating adhesion to urinary tract cell lines. In all, this work establishes InvL as an adhesin important for Acinetobacter's urinary tract virulence and represents the first report of a type II secretion system effector belonging to the intimin-invasin family.
Keywords: Acinetobacter; adhesin; infection; invasin; pathogenesis; type II secretion system; urinary tract infection; virulence.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
