

The host exocyst complex is targeted by a conserved bacterial type-III effector that promotes virulence
- PMID: 35640532
- PMCID: PMC9421483
- DOI: 10.1093/plcell/koac162
The host exocyst complex is targeted by a conserved bacterial type-III effector that promotes virulence
Abstract
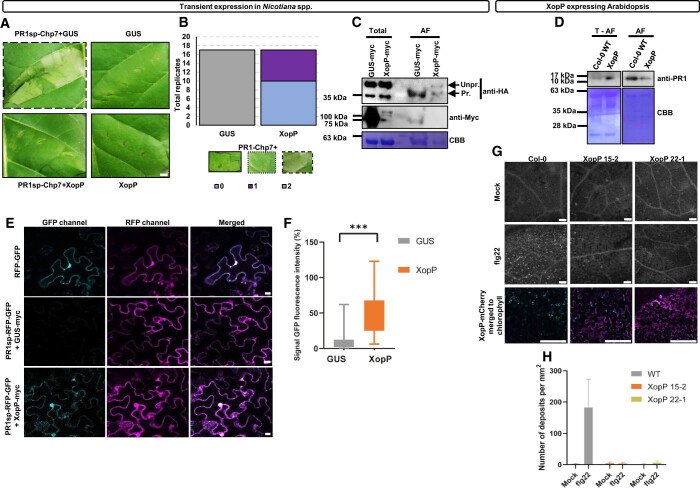
For most Gram-negative bacteria, pathogenicity largely depends on the type-III secretion system that delivers virulence effectors into eukaryotic host cells. The subcellular targets for the majority of these effectors remain unknown. Xanthomonas campestris, the causal agent of black rot disease of crucifers such as Brassica spp., radish, and turnip, delivers XopP, a highly conserved core-effector protein produced by X. campestris, which is essential for virulence. Here, we show that XopP inhibits the function of the host-plant exocyst complex by direct targeting of Exo70B, a subunit of the exocyst complex, which plays a significant role in plant immunity. XopP interferes with exocyst-dependent exocytosis and can do this without activating a plant NOD-like receptor that guards Exo70B in Arabidopsis. In this way, Xanthomonas efficiently inhibits the host's pathogen-associated molecular pattern (PAMP)-triggered immunity by blocking exocytosis of pathogenesis-related protein-1A, callose deposition, and localization of the FLAGELLIN SENSITIVE2 (FLS2) immune receptor to the plasma membrane, thus promoting successful infection. Inhibition of exocyst function without activating the related defenses represents an effective virulence strategy, indicating the ability of pathogens to adapt to host defenses by avoiding host immunity responses.
© American Society of Plant Biologists 2022. All rights reserved. For permissions, please email: journals.permissions@oup.com.
Figures








Comment in
-
Xanthomonas counteracts host immunity by targeting the exocyst complex.Plant Cell. 2022 Aug 25;34(9):3166-3167. doi: 10.1093/plcell/koac173. Plant Cell. 2022. PMID: 35762457 Free PMC article. No abstract available.
References
-
- Azevedo C, Santos-Rosa MJ, Shirasu K (2001) The U-box protein family in plants. Trends Plant Sci 6: 354–358 - PubMed
-
- Badel JL, Shimizu R, Oh HS, Collmer A (2006) A Pseudomonas syringae pv. tomato avrE1/hopM1 mutant is severely reduced in growth and lesion formation in tomato. Mol Plant-Microbe Interact 19: 99–111 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
