

Cryo-EM structures of human A2ML1 elucidate the protease-inhibitory mechanism of the A2M family
- PMID: 35641520
- PMCID: PMC9156758
- DOI: 10.1038/s41467-022-30758-x
Cryo-EM structures of human A2ML1 elucidate the protease-inhibitory mechanism of the A2M family
Abstract
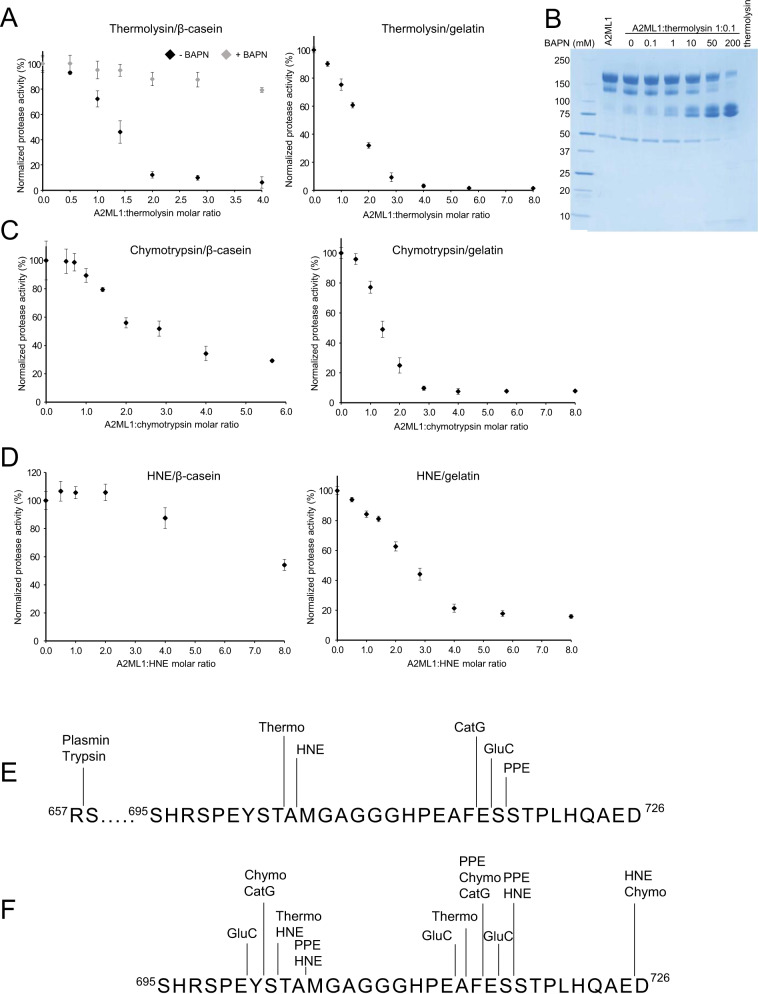
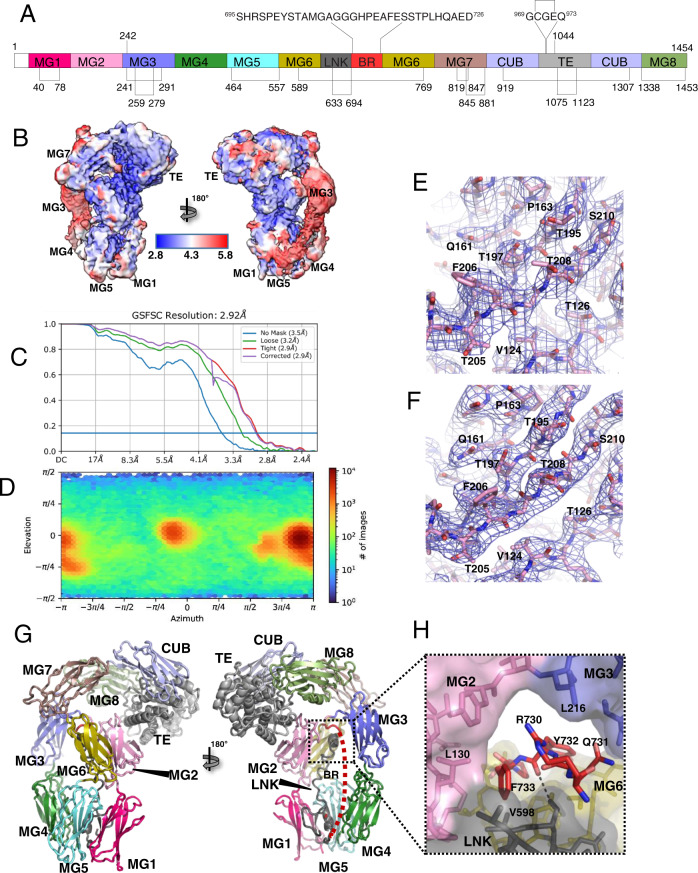
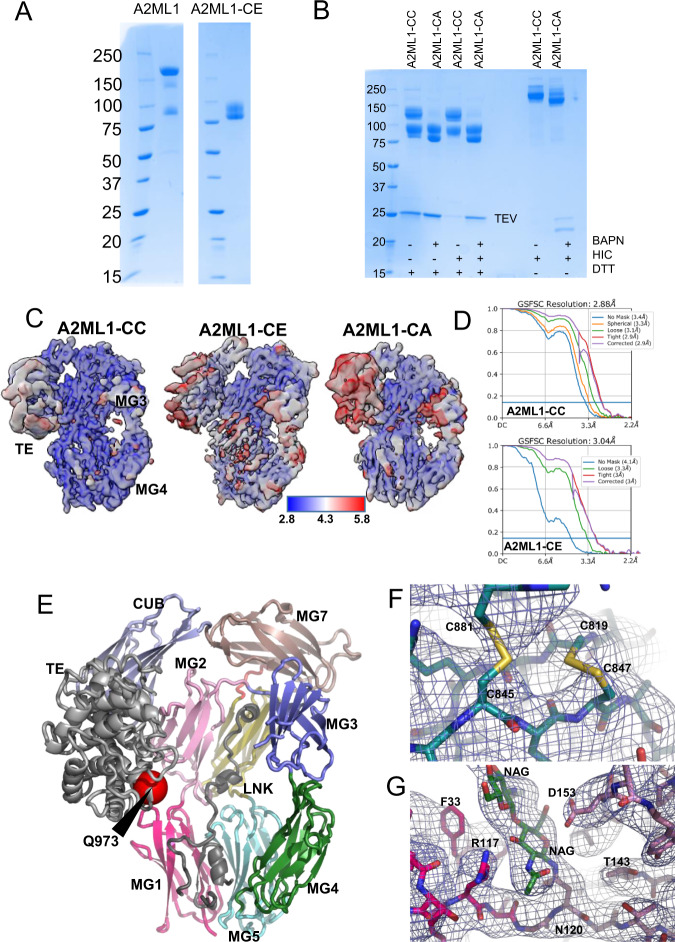
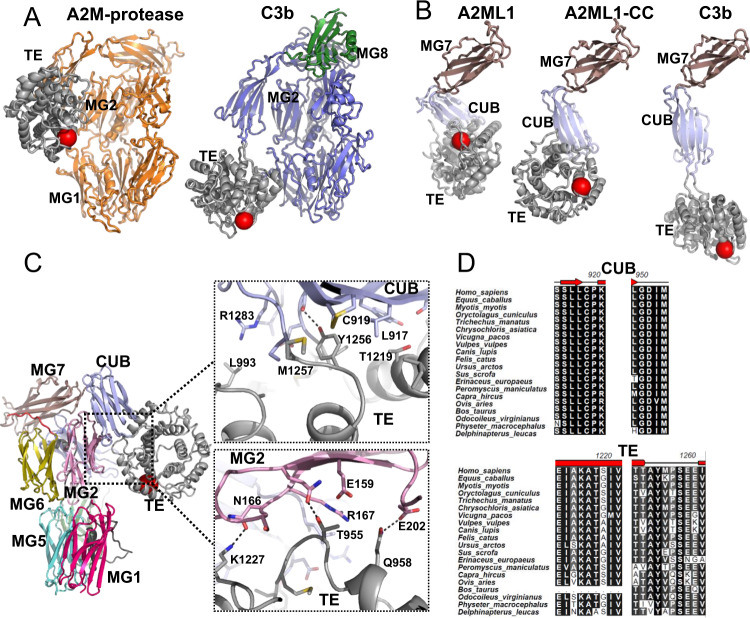
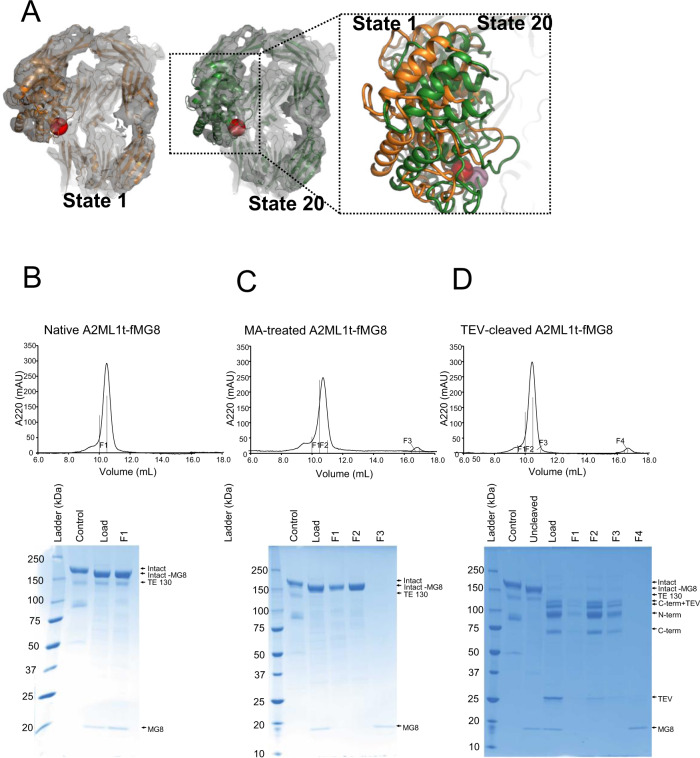
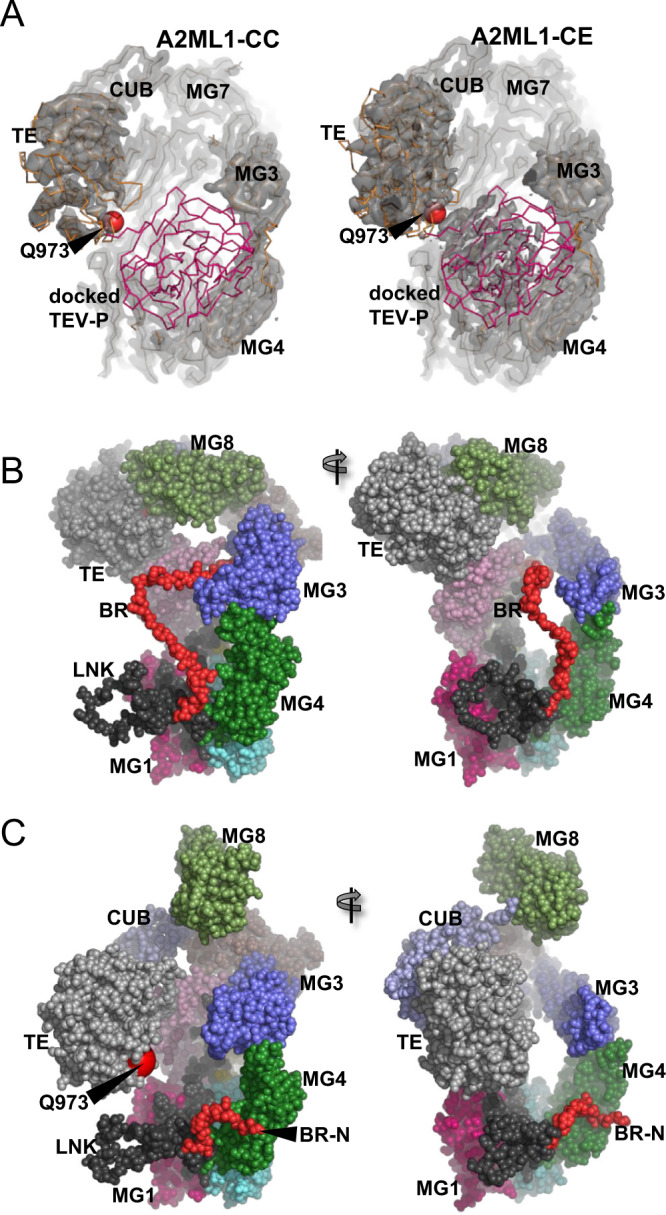
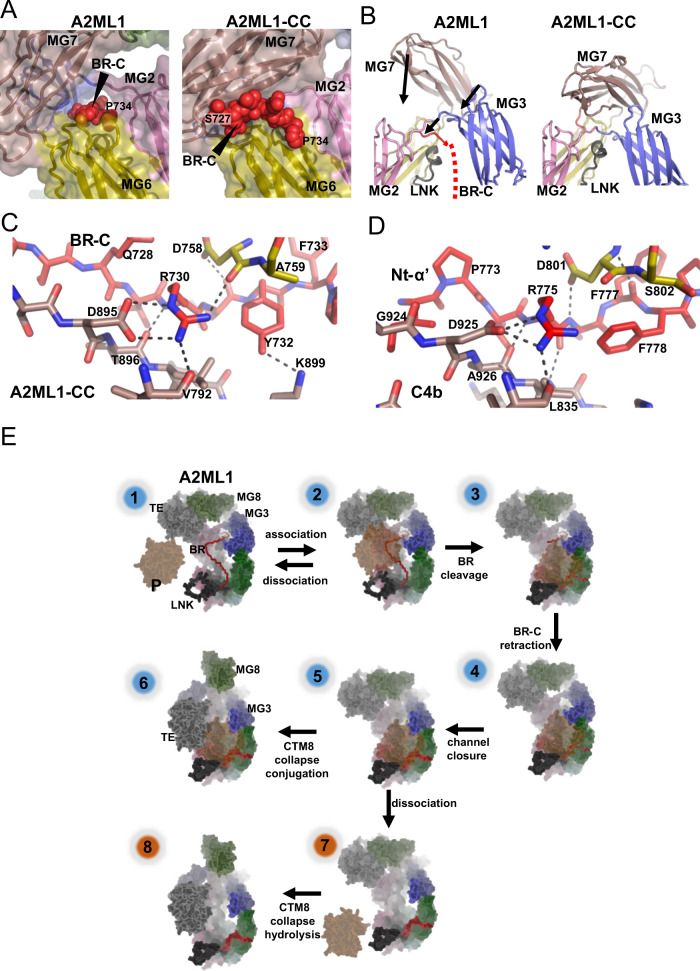
A2ML1 is a monomeric protease inhibitor belonging to the A2M superfamily of protease inhibitors and complement factors. Here, we investigate the protease-inhibitory mechanism of human A2ML1 and determine the structures of its native and protease-cleaved conformations. The functional inhibitory unit of A2ML1 is a monomer that depends on covalent binding of the protease (mediated by A2ML1's thioester) to achieve inhibition. In contrast to the A2M tetramer which traps proteases in two internal chambers formed by four subunits, in protease-cleaved monomeric A2ML1 disordered regions surround the trapped protease and may prevent substrate access. In native A2ML1, the bait region is threaded through a hydrophobic channel, suggesting that disruption of this arrangement by bait region cleavage triggers the extensive conformational changes that result in protease inhibition. Structural comparisons with complement C3/C4 suggest that the A2M superfamily of proteins share this mechanism for the triggering of conformational change occurring upon proteolytic activation.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
α2-Macroglobulin-like protein 1 can conjugate and inhibit proteases through their hydroxyl groups, because of an enhanced reactivity of its thiol ester.J Biol Chem. 2020 Dec 4;295(49):16732-16742. doi: 10.1074/jbc.RA120.015694. Epub 2020 Sep 25. J Biol Chem. 2020. PMID: 32978260 Free PMC article.
-
The conformational change of the protease inhibitor α2-macroglobulin is triggered by the retraction of the cleaved bait region from a central channel.J Biol Chem. 2022 Aug;298(8):102230. doi: 10.1016/j.jbc.2022.102230. Epub 2022 Jul 1. J Biol Chem. 2022. PMID: 35787371 Free PMC article.
-
Structural Investigations of Human A2M Identify a Hollow Native Conformation That Underlies Its Distinctive Protease-Trapping Mechanism.Mol Cell Proteomics. 2021;20:100090. doi: 10.1016/j.mcpro.2021.100090. Epub 2021 May 6. Mol Cell Proteomics. 2021. PMID: 33964423 Free PMC article.
-
Structural and functional insight into pan-endopeptidase inhibition by α2-macroglobulins.Biol Chem. 2017 Aug 28;398(9):975-994. doi: 10.1515/hsz-2016-0329. Biol Chem. 2017. PMID: 28253193 Review.
-
The phylogeny and evolution of the thioester bond-containing proteins C3, C4 and alpha 2-macroglobulin.Immunol Rev. 1998 Dec;166:15-26. doi: 10.1111/j.1600-065x.1998.tb01249.x. Immunol Rev. 1998. PMID: 9914899 Review.
Cited by
-
Engineering New Protease Inhibitors Using α2-Macroglobulin.Methods Mol Biol. 2024;2747:279-294. doi: 10.1007/978-1-0716-3589-6_21. Methods Mol Biol. 2024. PMID: 38038947
-
Audiologic Measures in an Indigenous Community with A2ML1- and FUT2-Related Otitis Media.Genet Test Mol Biomarkers. 2023 Jan;27(1):12-17. doi: 10.1089/gtmb.2022.0171. Genet Test Mol Biomarkers. 2023. PMID: 36719978 Free PMC article.
-
Cryo-EM analysis of complement C3 reveals a reversible major opening of the macroglobulin ring.Nat Struct Mol Biol. 2025 May;32(5):884-895. doi: 10.1038/s41594-024-01467-4. Epub 2025 Jan 23. Nat Struct Mol Biol. 2025. PMID: 39849196
-
Multi-omics integration and machine learning uncover molecular basal-like subtype of pancreatic cancer and implicate A2ML1 in promoting tumor epithelial-mesenchymal transition.J Transl Med. 2025 Jul 4;23(1):741. doi: 10.1186/s12967-025-06711-z. J Transl Med. 2025. PMID: 40615919 Free PMC article.
-
Recent cryogenic electron microscopy structures of human A2M may not be representative of the native protein.Proc Natl Acad Sci U S A. 2022 Sep 13;119(37):e2210218119. doi: 10.1073/pnas.2210218119. Epub 2022 Aug 16. Proc Natl Acad Sci U S A. 2022. PMID: 35972981 Free PMC article. No abstract available.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
