

Epigenetic regulation of BAF60A determines efficiency of miniature swine iPSC generation
- PMID: 35641537
- PMCID: PMC9156668
- DOI: 10.1038/s41598-022-12919-6
Epigenetic regulation of BAF60A determines efficiency of miniature swine iPSC generation
Erratum in
-
Author Correction: Epigenetic regulation of BAF60A determines efficiency of miniature swine iPSC generation.Sci Rep. 2022 Jun 17;12(1):10193. doi: 10.1038/s41598-022-14878-4. Sci Rep. 2022. PMID: 35715616 Free PMC article. No abstract available.
Abstract
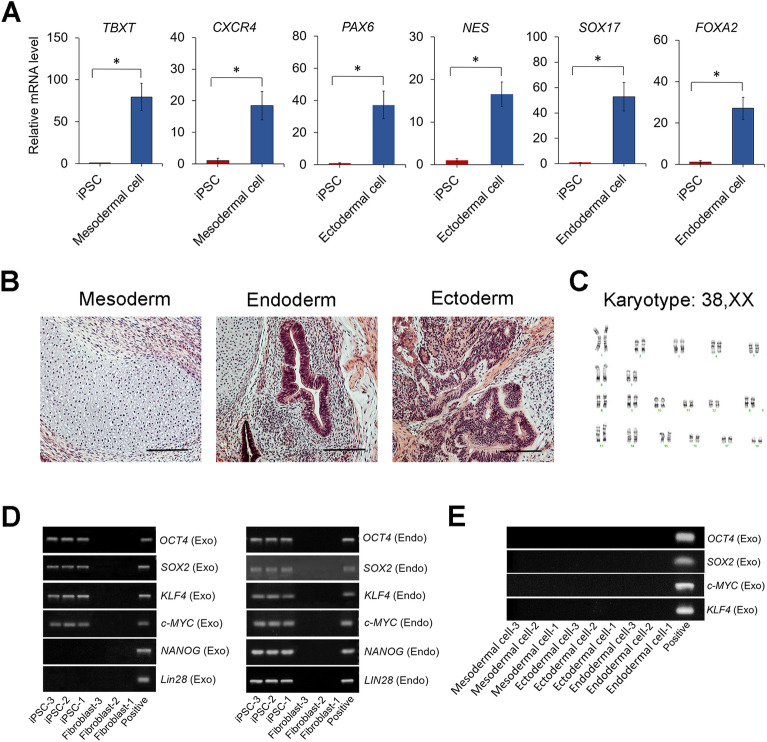
Miniature pigs are an ideal animal model for translational research to evaluate stem cell therapies and regenerative applications. While the derivation of induced pluripotent stem cells (iPSCs) from miniature pigs has been demonstrated, there is still a lack of a reliable method to generate and maintain miniature pig iPSCs. In this study, we derived iPSCs from fibroblasts of Wisconsin miniature swine (WMS), Yucatan miniature swine (YMS), and Göttingen minipigs (GM) using our culture medium. By comparing cells of the different pig breeds, we found that YMS fibroblasts were more efficiently reprogrammed into iPSCs, forming colonies with well-defined borders, than WMS and GM fibroblasts. We also demonstrated that YMS iPSC lines with a normal pig karyotype gave rise to cells of the three germ layers in vitro and in vivo. Mesenchymal stromal cells expressing phenotypic characteristics were derived from established iPSC lines as an example of potential applications. In addition, we found that the expression level of the switch/sucrose nonfermentable component BAF60A regulated by STAT3 signaling determined the efficiency of pig iPSC generation. The findings of this study provide insight into the underlying mechanism controlling the reprogramming efficiency of miniature pig cells to develop a viable strategy to enhance the generation of iPSCs for biomedical research.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures






References
-
- Barone AAL, et al. Genetically modified porcine split-thickness skin grafts as an alternative to allograft for provision of temporary wound coverage: Preliminary characterization. Burns. 2015;41(3):565–574. - PubMed
-
- Summerfield A, Meurens F, Ricklin ME. The immunology of the porcine skin and its value as a model for human skin. Mol. Immunol. 2015;66(1):14–21. - PubMed
-
- Lai L, et al. Production of α-1,3-galactosyltransferase knockout pigs by nuclear transfer cloning. Science. 2002;295(5557):1089–1092. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
