

Nutritional immunity: the battle for nutrient metals at the host-pathogen interface
- PMID: 35641670
- PMCID: PMC9153222
- DOI: 10.1038/s41579-022-00745-6
Nutritional immunity: the battle for nutrient metals at the host-pathogen interface
Abstract
Trace metals are essential micronutrients required for survival across all kingdoms of life. From bacteria to animals, metals have critical roles as both structural and catalytic cofactors for an estimated third of the proteome, representing a major contributor to the maintenance of cellular homeostasis. The reactivity of metal ions engenders them with the ability to promote enzyme catalysis and stabilize reaction intermediates. However, these properties render metals toxic at high concentrations and, therefore, metal levels must be tightly regulated. Having evolved in close association with bacteria, vertebrate hosts have developed numerous strategies of metal limitation and intoxication that prevent bacterial proliferation, a process termed nutritional immunity. In turn, bacterial pathogens have evolved adaptive mechanisms to survive in conditions of metal depletion or excess. In this Review, we discuss mechanisms by which nutrient metals shape the interactions between bacterial pathogens and animal hosts. We explore the cell-specific and tissue-specific roles of distinct trace metals in shaping bacterial infections, as well as implications for future research and new therapeutic development.
© 2022. Springer Nature Limited.
Conflict of interest statement
The authors declare no competing interests.
Figures
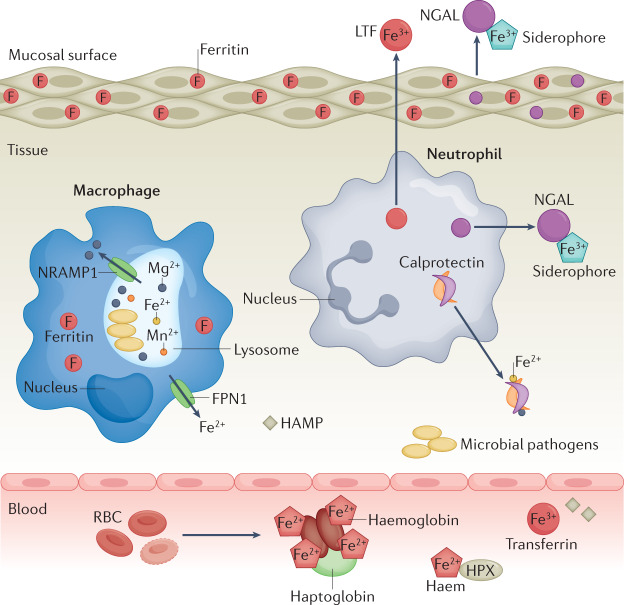
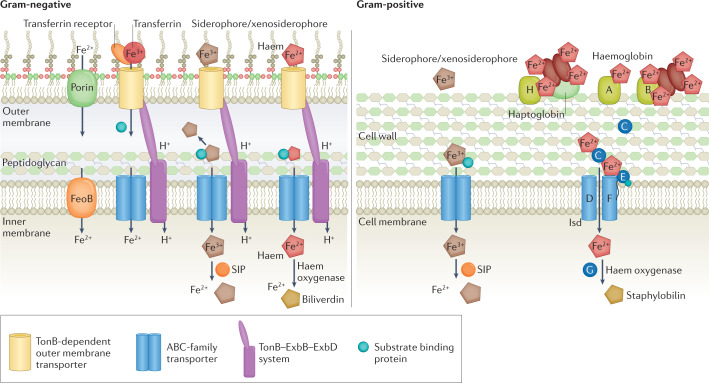
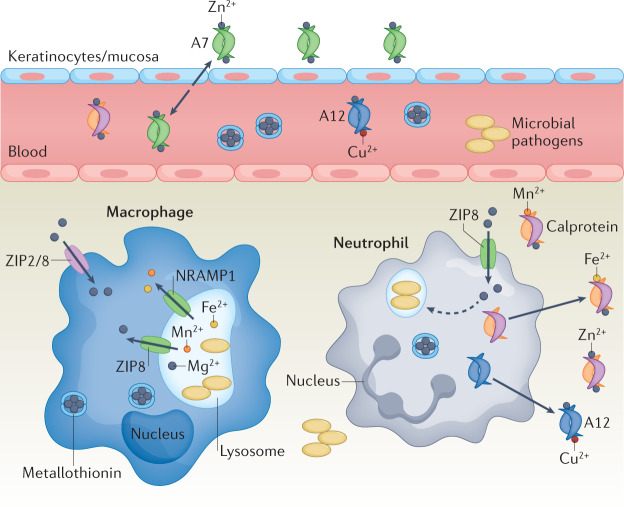
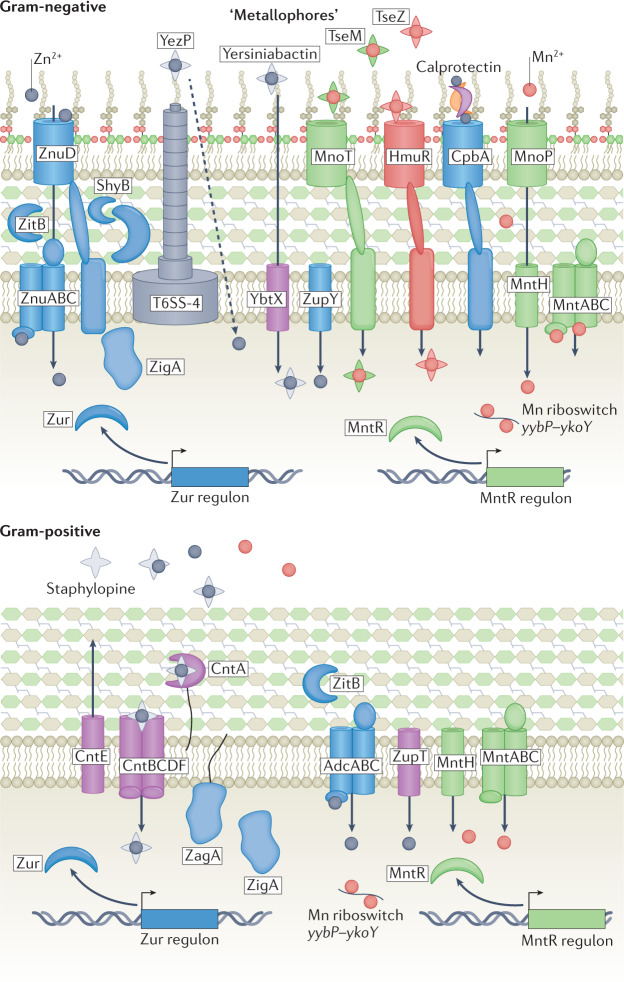
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
