An official website of the United States government
The .gov means it’s official.
Federal government websites often end in .gov or .mil. Before
sharing sensitive information, make sure you’re on a federal
government site.
The site is secure.
The https:// ensures that you are connecting to the
official website and that any information you provide is encrypted
and transmitted securely.
The discovery of microRNAs and their role in diseases was a breakthrough that inspired research into microRNAs as drug targets. Cardiovascular diseases are an area in which limitations of conventional pharmacotherapy are highly apparent and where microRNA-based drugs have appreciably progressed into preclinical and clinical testing. In this Review, we summarize the current state of microRNAs as therapeutic targets in the cardiovascular system. We report recent advances in the identification and characterization of microRNAs, their manipulation and clinical translation, and discuss challenges and perspectives toward clinical application.
Conflict of interest: SE is an inventor on patents related to miRNA-based therapeutics filed by the University of Würzburg and the Technical University of Munich (US8592389B2, WO 2021/205032 A1), and is founder of rnatics GmbH.
Figures
Figure 1. MicroRNA life cycle, and identification…
Figure 1. MicroRNA life cycle, and identification of microRNAs and their targets.
( A )…
Figure 1. MicroRNA life cycle, and identification of microRNAs and their targets.
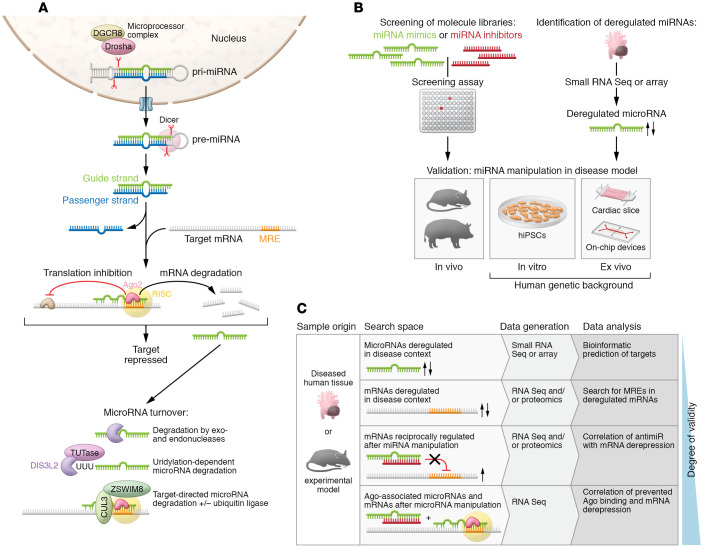
(A) Canonical processing, functional activation, mechanism of action, and degradation pathways of microRNAs. Canonical microRNA biogenesis starts from larger hairpin RNA molecules (pri-miRNAs), which are generated by RNA Pol II transcription of microRNA genes or clusters, or which occur as part of introns. A microprocessor complex that contains the endonuclease Drosha, the DiGeorge critical region 8 protein (DGCR8), and other factors then cleaves these pri-miRNAs. The resulting pre-miRNA is exported to the cytoplasm, where the nuclease Dicer tailors it to 21 to 22 nucleotides in length. There are also noncanonical mechanisms of microRNA biogenesis, some of which bypass the microprocessor complex or Dicer. After processing to a duplex of 21–22 nucleotides in length each, one strand, termed the guide strand, becomes part of the RNA-induced silencing complex (RISC), whereas the passenger strand (or *-strand) undergoes accelerated degradation. If both strands are maintained, they can adopt individual functions, as demonstrated for cardiovascular miR-21 and miR-126 (11, 12). Another exception are microRNA strands that localize to the nucleus, where they function in unusual manners (12, 13). Degradation of microRNAs involves exonucleases XRN-1, PNPase old-35, and RRP41 (17) or the endonuclease Tudor-SN (154). The nuclease DIS3L2 degrades a subset of microRNAs after modification by terminal uridyltransferases (TUTases) (155). Mechanisms of target-directed microRNA degradation (TDMD) have been resolved, including the involvement of ubiquitin ligases (25, 26). (B) Routes toward the identification and validation of disease-relevant cardiovascular microRNAs. (C) Approaches for the identification of microRNA targets.
Figure 2. Functions of microRNAs in the…
Figure 2. Functions of microRNAs in the cardiovascular system.
( A ) Table summarizing microRNA…
Figure 2. Functions of microRNAs in the cardiovascular system.
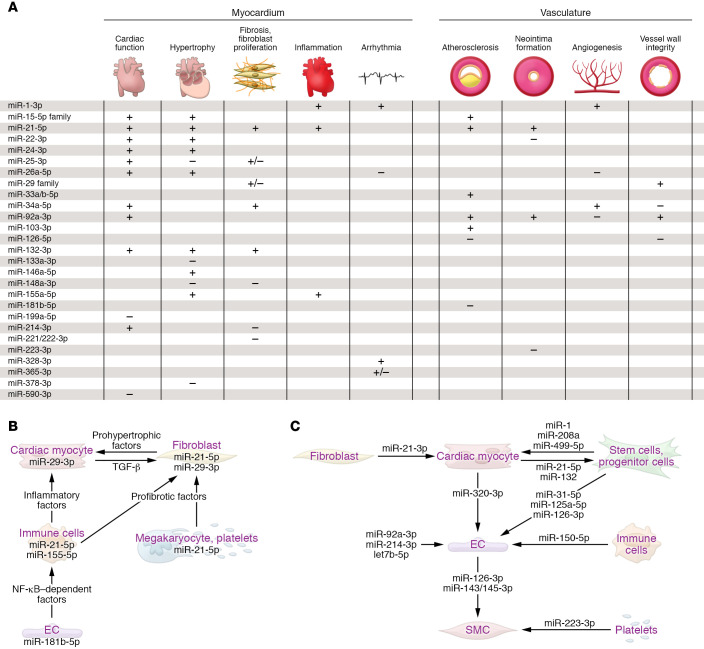
(A) Table summarizing microRNA functions in myocardium and vasculature. + indicates that the process is promoted by the indicated microRNA, – indicates the pathophysiologic process is prevented by the indicated microRNA. Information on microRNAs that promote or impair cardiac function, after their elevation or inhibition, is provided in the respective column. (B) Exemplary microRNAs that control targets involved in cell-to-cell communication in the cardiovascular system (information compiled from studies cited in Table 1). (C) Paracrine roles of exemplary secreted microRNAs in the cardiovascular system. Atheroprotective effects are exerted by extracellular miR-126-3p (184) and miR-143-3p/miR-145-3p, proangiogenic effects are exerted by exosomal miR-143-3p, miR-222-5p, miR-92a-3p, and miR-214-3p, whereas miR-320-3p confers the opposite effect. The passenger (3′) strand of miR-21 is enriched in exosomes from cardiac fibroblasts, promoting cardiac myocyte hypertrophy (156), whereas the miR-21 guide strand, released by endometrial mesenchymal stem cells, is cardioprotective by promoting cell survival and angiogenesis (157). In the retrograde direction, several microRNAs of myocardial origin promote the mobilization of progenitor cells in bone marrow (102). miR-223-3p is delivered by platelets and regulates differentiation and proliferation of vascular SMCs (193). For an overview on these and other cardiovascular microRNAs with proposed paracrine function, see refs. –. EC, endothelial cell; SMC, smooth muscle cell.
Figure 3. Molecular vehicles for microRNA modulators…
Figure 3. Molecular vehicles for microRNA modulators and their functionalization.
Improved nuclease resistance by the…
Figure 3. Molecular vehicles for microRNA modulators and their functionalization.
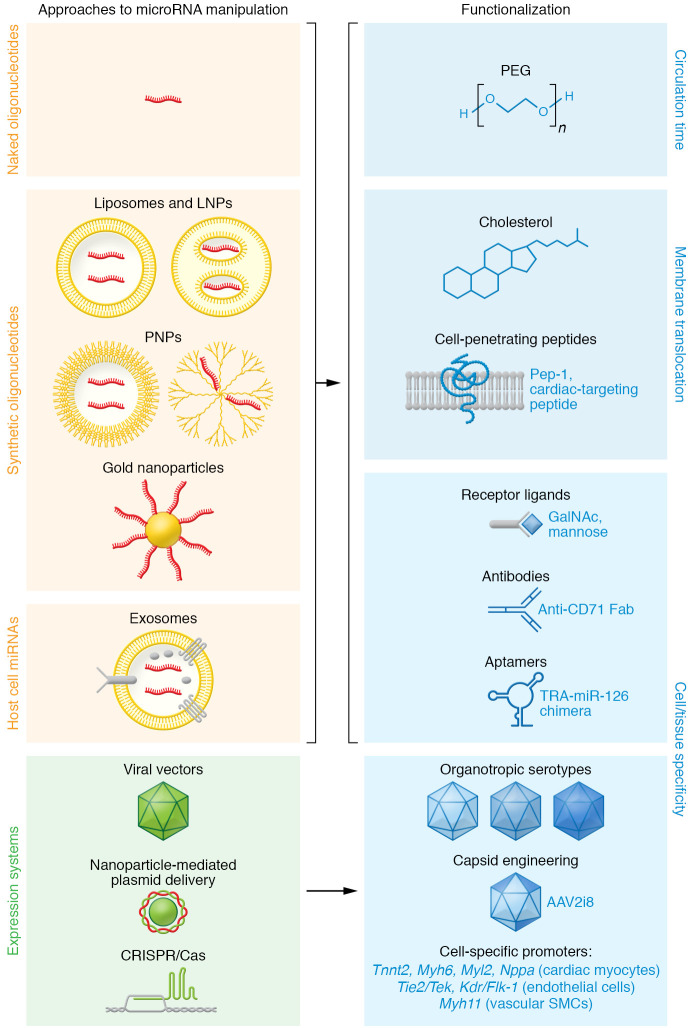
Improved nuclease resistance by the use of modified nucleotides in synthetic oligonucleotides allows for application as “naked” molecules (112). Their embedding in liposomes or lipid nanoparticles (LNPs) or polymer-based nanoparticles (PNPs) can improve cell entry via endocytosis (113). Metal particles such as gold have been used as carriers for oligonucleotides (113) and plasmids (126). Exosomes with microRNA cargo can be isolated from native sources or engineered for optimized microRNA loading or cell specificity (104, 105). Oligonucleotides or their carriers can be further functionalized by conjugation to improve their circulation time (e.g., by PEGylation), membrane penetrance (e.g., cholesterol, cell-penetrating peptides), or to enhance their cell- or tissue-specific delivery (e.g., by coupling to receptor ligands, antibody fragments, or aptamers). TRA, transferrin receptor aptamer. Viral vectors and their organotropic serotypes, particularly adeno-associated virus (AAV), can be utilized for the expression or genetic inactivation (e.g., using CRISPR/Cas systems) of microRNAs or their targets. Improved transduction and/or tropism can be achieved by engineering AAVs (122, 123), and the use of cell-type-specific promoters adds further improvement. Exemplary promoters are denoted for gene expression in cardiac myocytes (Tnnt2, cardiac troponin T2; Myh6, myosin heavy chain 6; Myl2, myosin light chain 2; Nppa, natriuretic peptide A), in endothelial cells (protein tyrosine kinase Tie2/Tek; Kdr/Flk-1, kinase insert domain receptor/fetal liver kinase 1), and in vascular smooth muscle cells (Myh11, myosin heavy chain 11). For a critical review on endothelial cell–specific promoters, see Chakraborty et al. (158).
Lee RC, et al. The C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14. Cell. 1993;75(5):843–854. doi: 10.1016/0092-8674(93)90529-Y.
-
DOI
-
PubMed
Wightman B, et al. Posttranscriptional regulation of the heterochronic gene lin-14 by lin-4 mediates temporal pattern formation in C. elegans. Cell. 1993;75(5):855–862. doi: 10.1016/0092-8674(93)90530-4.
-
DOI
-
PubMed
Friedman RC, et al. Most mammalian mRNAs are conserved targets of microRNAs. Genome Res. 2009;19(1):92–105. doi: 10.1101/gr.082701.108.
-
DOI
-
PMC
-
PubMed
Kozomara A, et al. miRBase: from microRNA sequences to function. Nucleic Acids Res. 2019;47(d1):D155–D162. doi: 10.1093/nar/gky1141.
-
DOI
-
PMC
-
PubMed
Fromm B, et al. MirGeneDB 2.0: the metazoan microRNA complement. Nucleic Acids Res. 2020;48(d1):132–141.
-
PMC
-
PubMed