

HIRA Supports Hepatitis B Virus Minichromosome Establishment and Transcriptional Activity in Infected Hepatocytes
- PMID: 35643233
- PMCID: PMC9304598
- DOI: 10.1016/j.jcmgh.2022.05.007
HIRA Supports Hepatitis B Virus Minichromosome Establishment and Transcriptional Activity in Infected Hepatocytes
Abstract
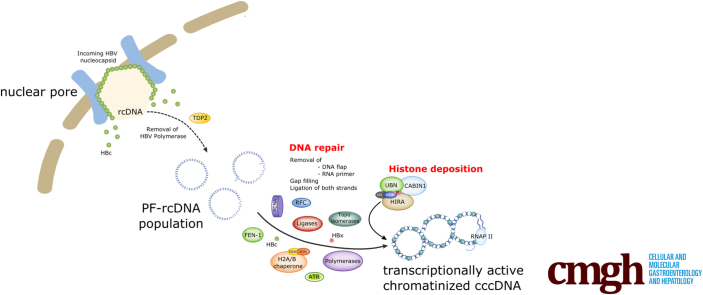
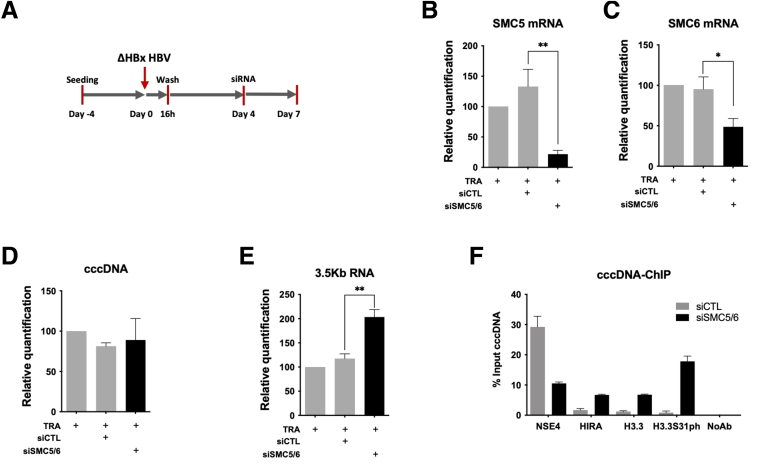
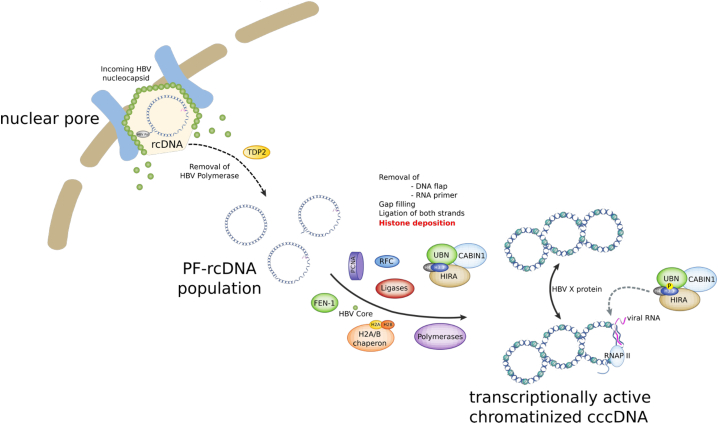
Background & aims: Upon hepatitis B virus (HBV) infection, partially double-stranded viral DNA converts into a covalently closed circular chromatinized episomal structure (cccDNA). This form represents the long-lived genomic reservoir responsible for viral persistence in the infected liver. Although the involvement of host cell DNA damage response in cccDNA formation has been established, this work investigated the yet-to-be-identified histone dynamics on cccDNA during early phases of infection in human hepatocytes.
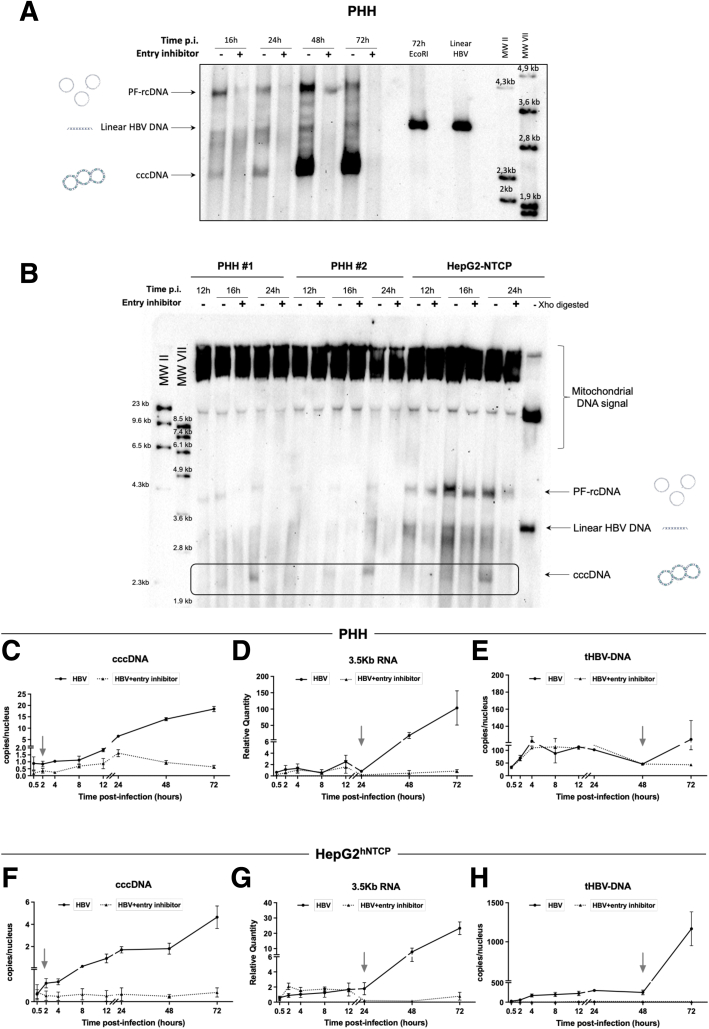
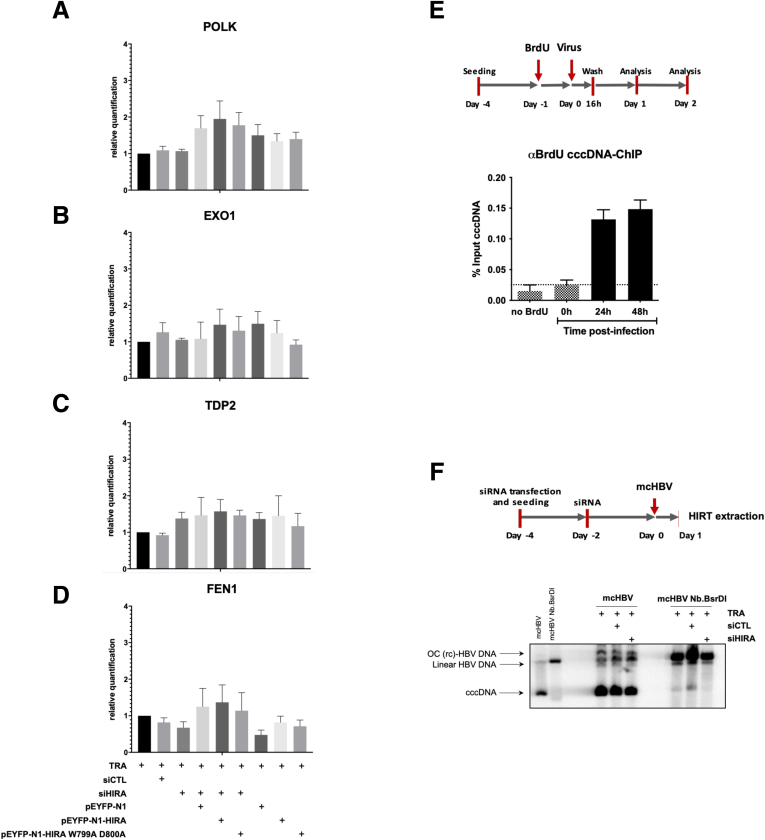
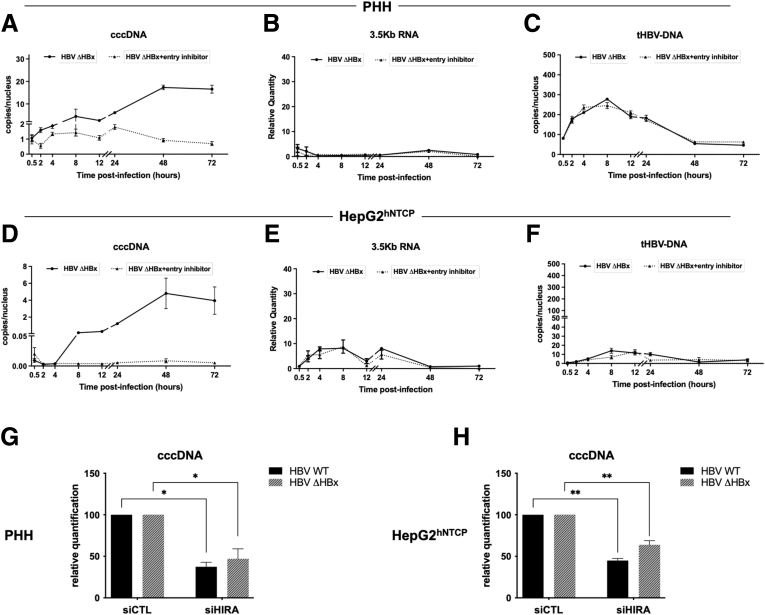
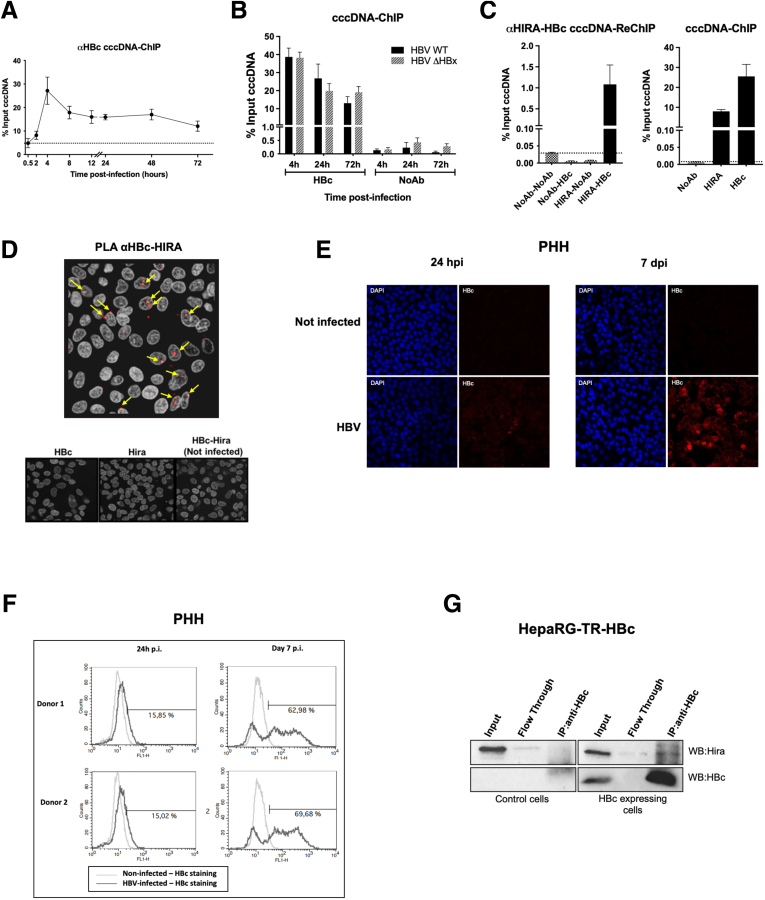
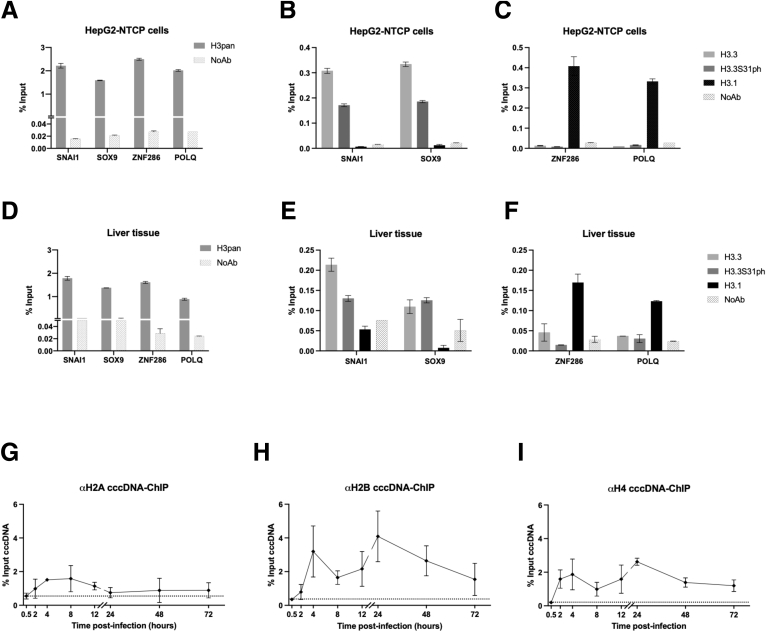
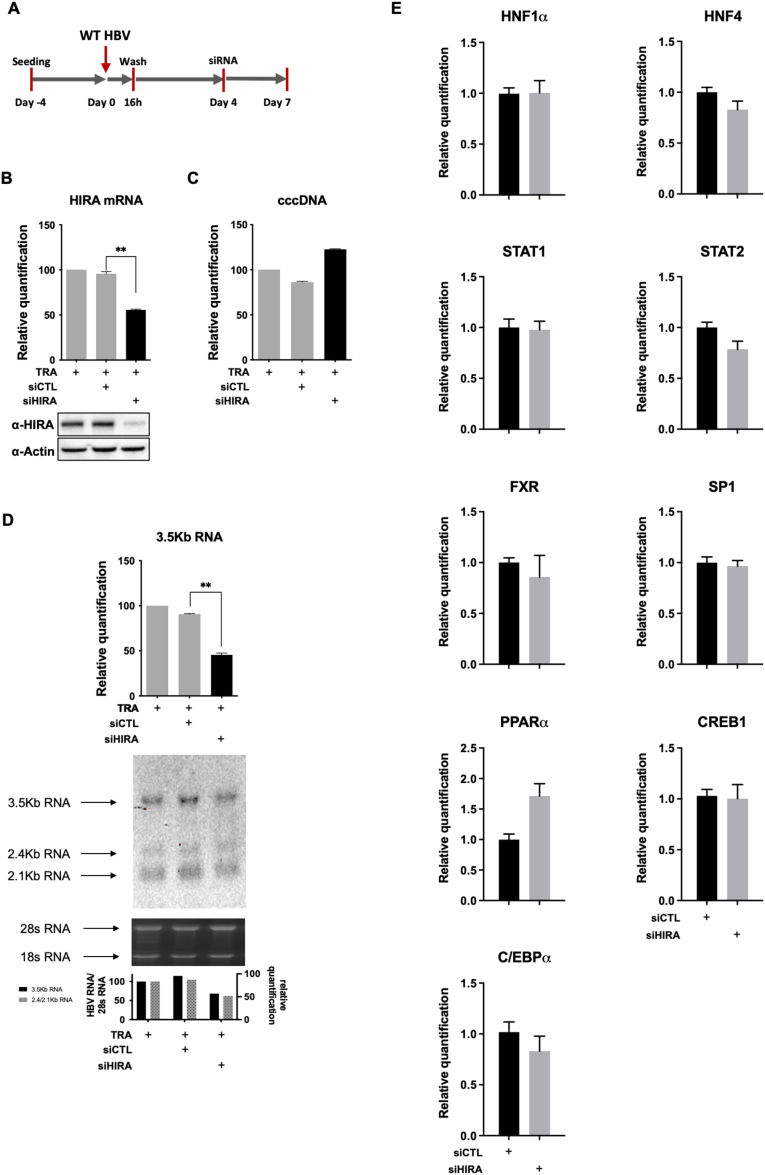
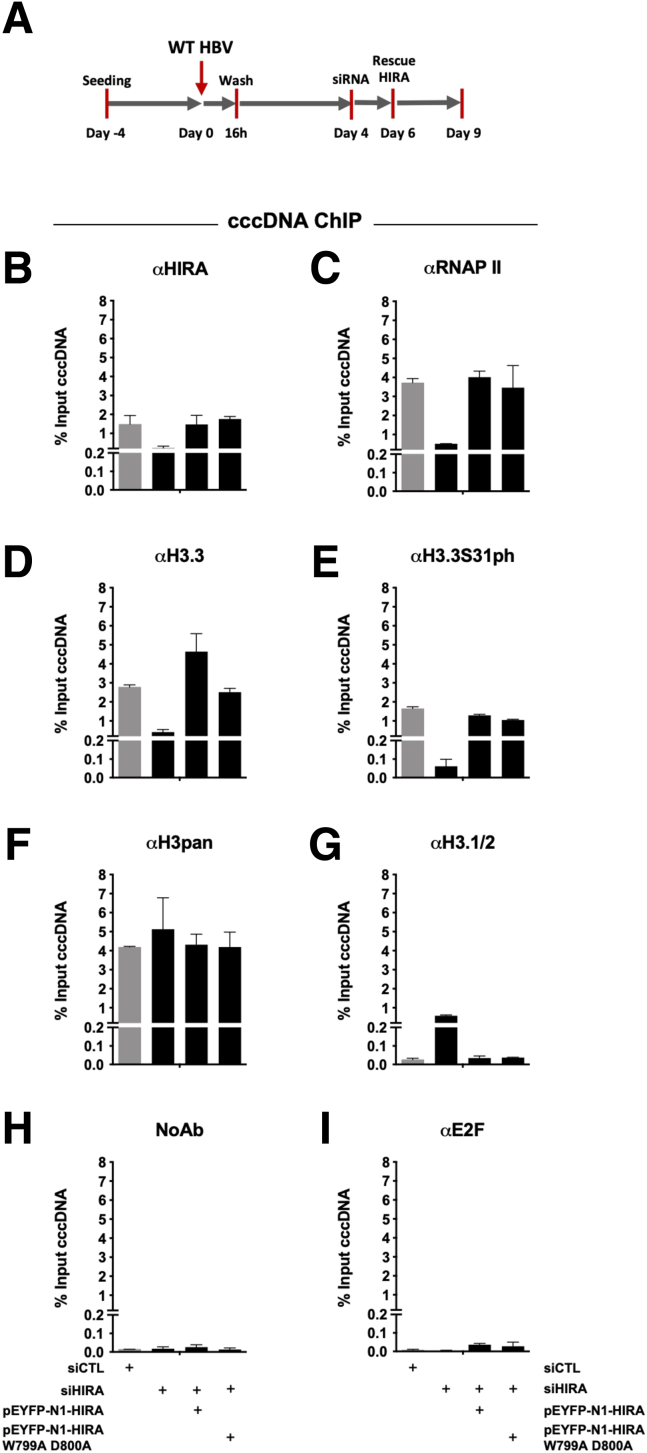
Methods: Detailed studies of host chromatin-associated factors were performed in cell culture models of natural infection (ie, Na+-taurocholate cotransporting polypeptide (NTCP)-overexpressing HepG2 cells, HepG2hNTCP) and primary human hepatocytes infected with HBV, by cccDNA-specific chromatin immunoprecipitation and loss-of-function experiments during early kinetics of viral minichromosome establishment and onset of viral transcription.
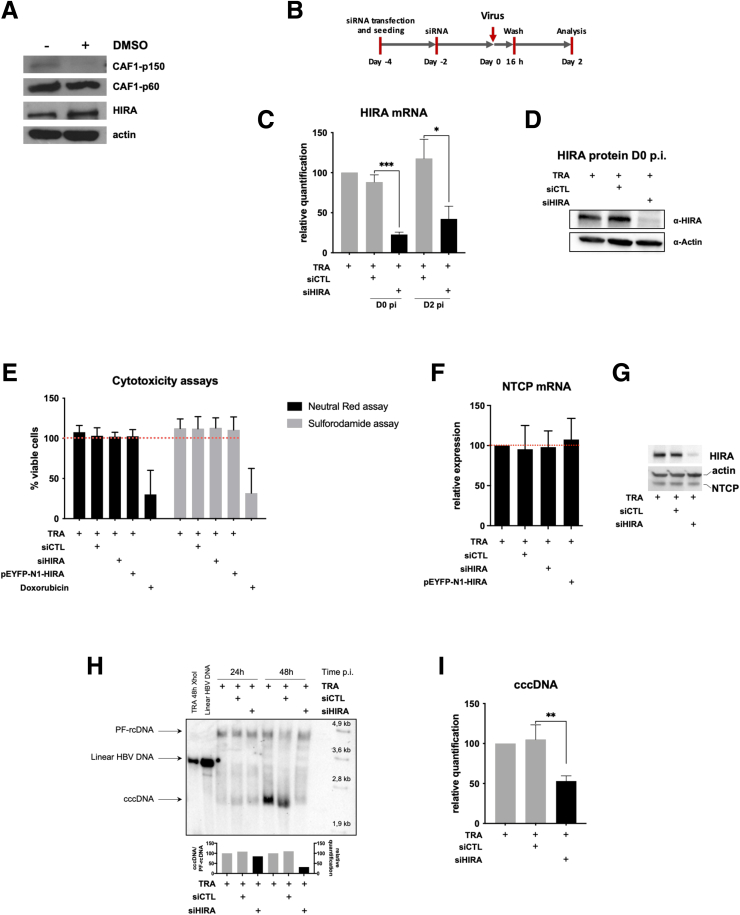
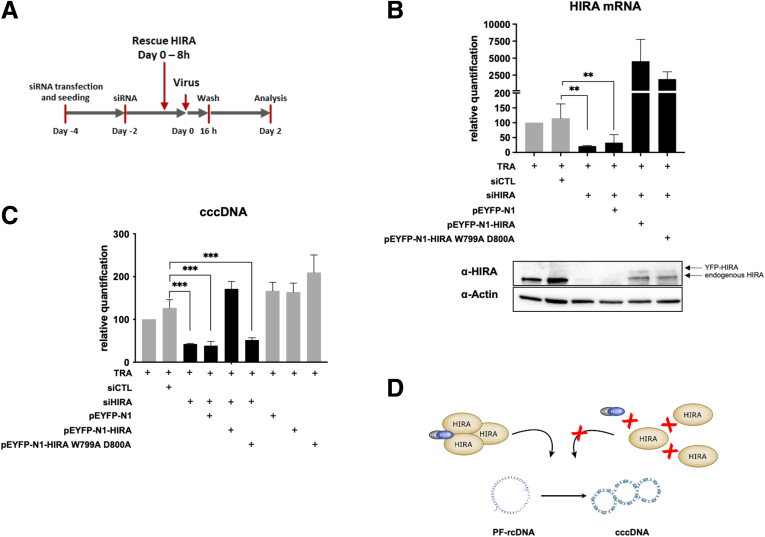
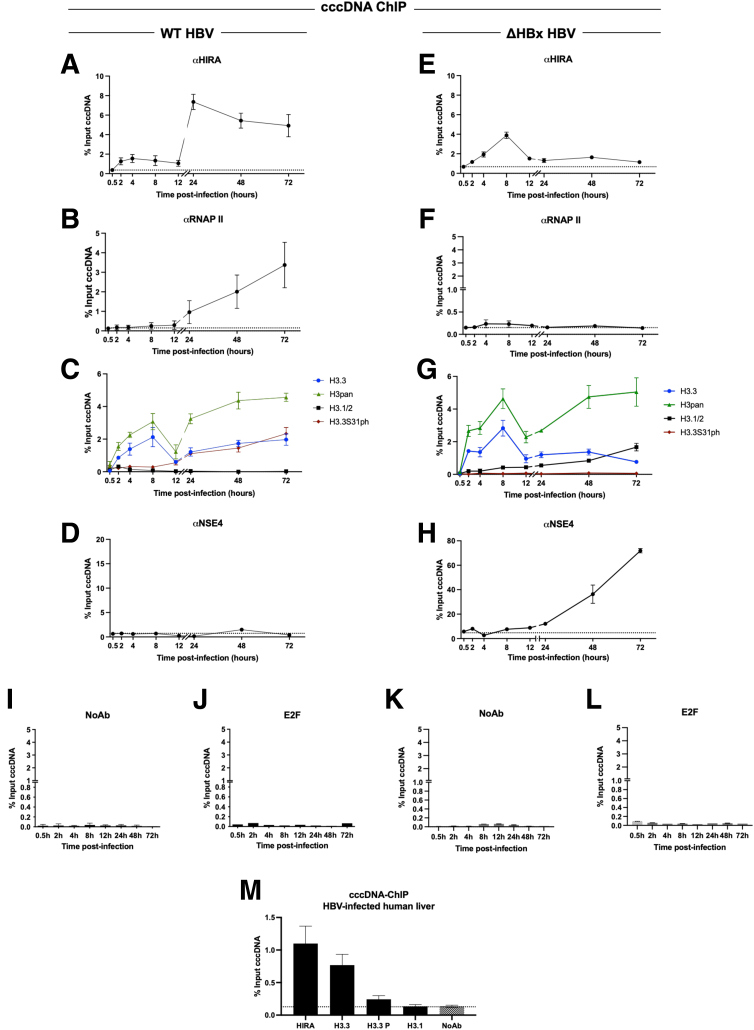
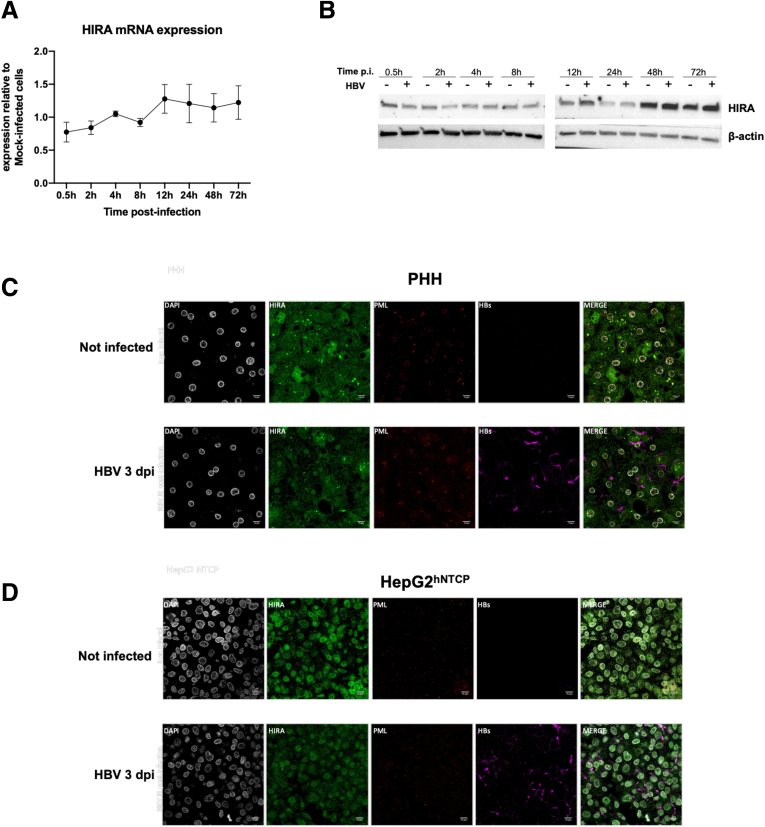
Results: Our results show that cccDNA formation requires the deposition of the histone variant H3.3 via the histone regulator A (HIRA)-dependent pathway. This occurs simultaneously with repair of the cccDNA precursor and independently from de novo viral protein expression. Moreover, H3.3 in its S31 phosphorylated form appears to be the preferential H3 variant found on transcriptionally active cccDNA in infected cultured cells and human livers. HIRA depletion after cccDNA pool establishment showed that HIRA recruitment is required for viral transcription and RNA production.
Conclusions: Altogether, we show a crucial role for HIRA in the interplay between HBV genome and host cellular machinery to ensure the formation and active transcription of the viral minichromosome in infected hepatocytes.
Keywords: H3.3; HBV; HIRA; cccDNA; chromatin.
Copyright © 2022 The Authors. Published by Elsevier Inc. All rights reserved.
Figures
Comment in
-
HIRA in Hepatitis B Virus Minichromosome Regulation: Another Piece of the Puzzle.Cell Mol Gastroenterol Hepatol. 2022;14(3):718-719. doi: 10.1016/j.jcmgh.2022.05.009. Epub 2022 Jun 9. Cell Mol Gastroenterol Hepatol. 2022. PMID: 35691338 Free PMC article. No abstract available.
