

TNF-α/IFN-γ synergy amplifies senescence-associated inflammation and SARS-CoV-2 receptor expression via hyper-activated JAK/STAT1
- PMID: 35645319
- PMCID: PMC9197409
- DOI: 10.1111/acel.13646
TNF-α/IFN-γ synergy amplifies senescence-associated inflammation and SARS-CoV-2 receptor expression via hyper-activated JAK/STAT1
Abstract
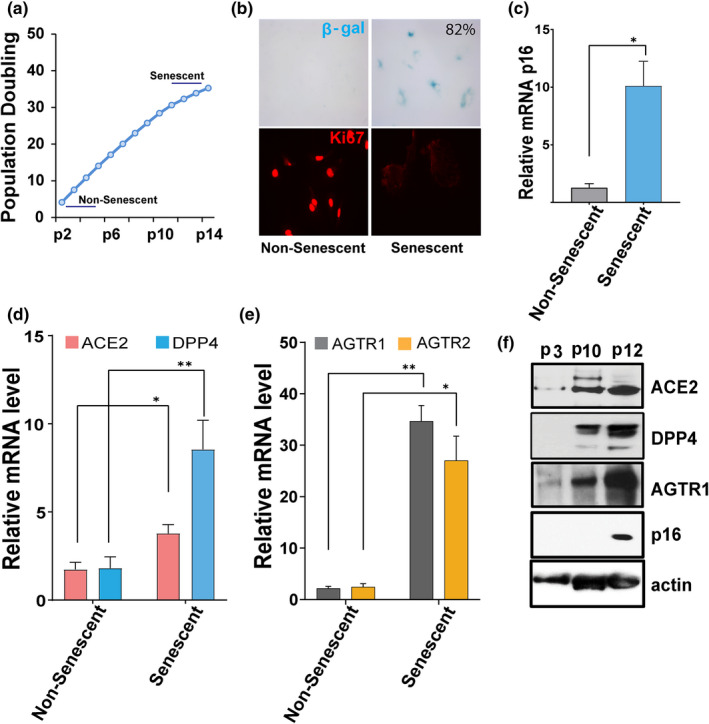
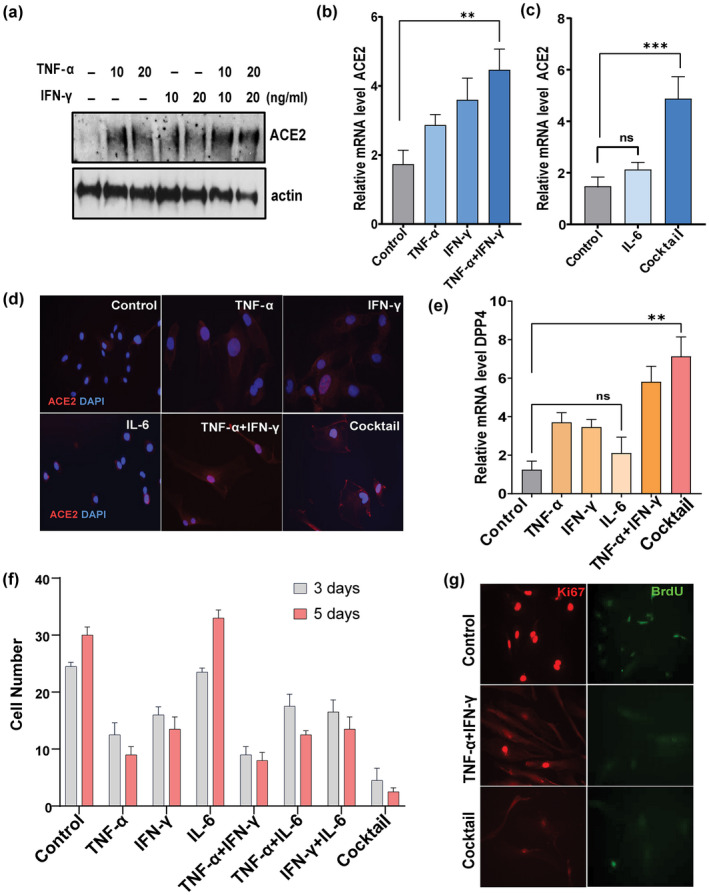
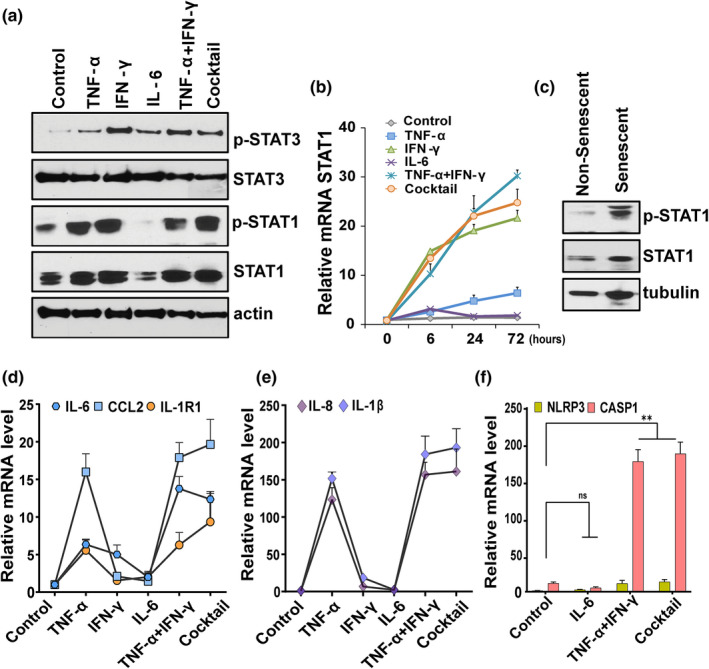
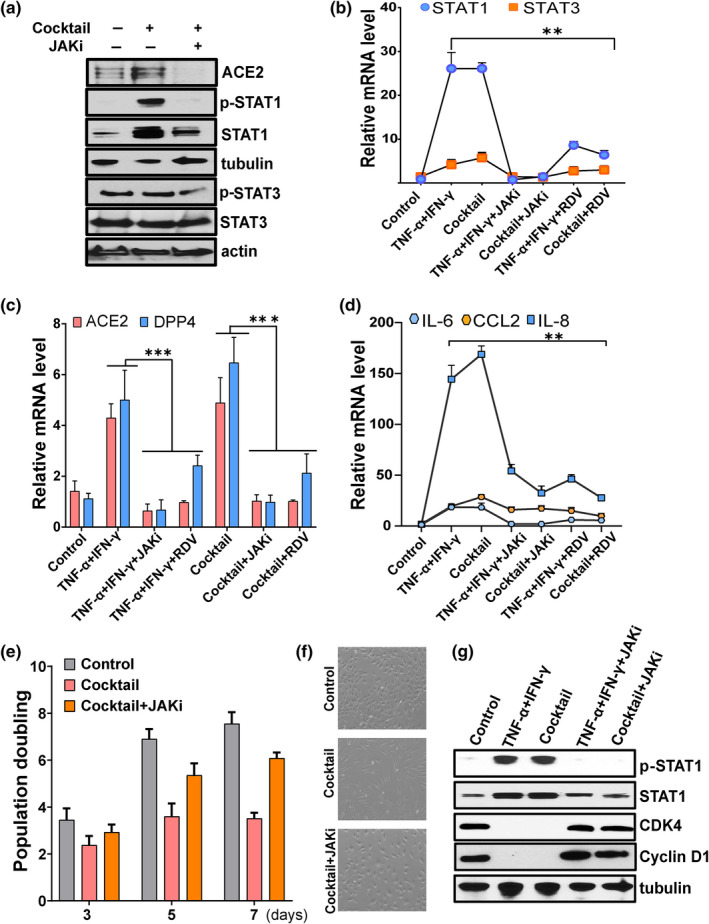
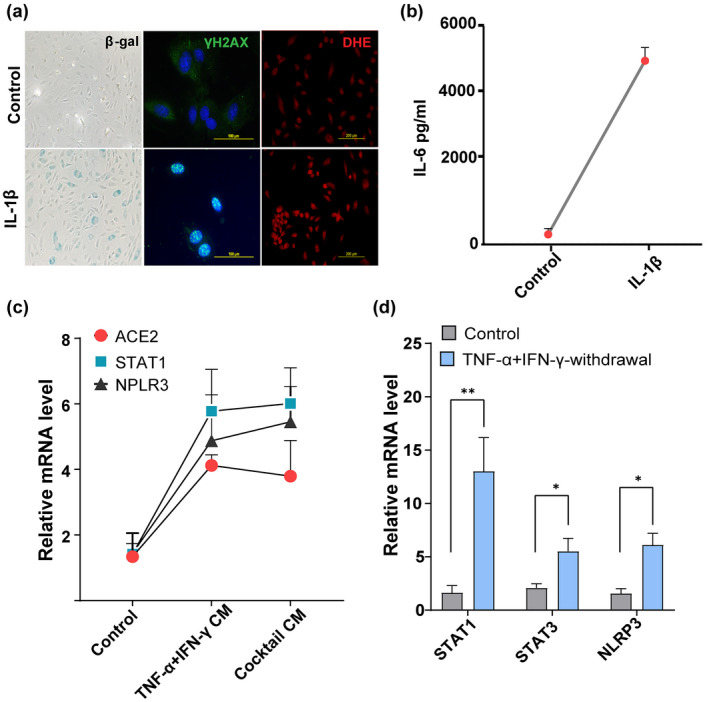
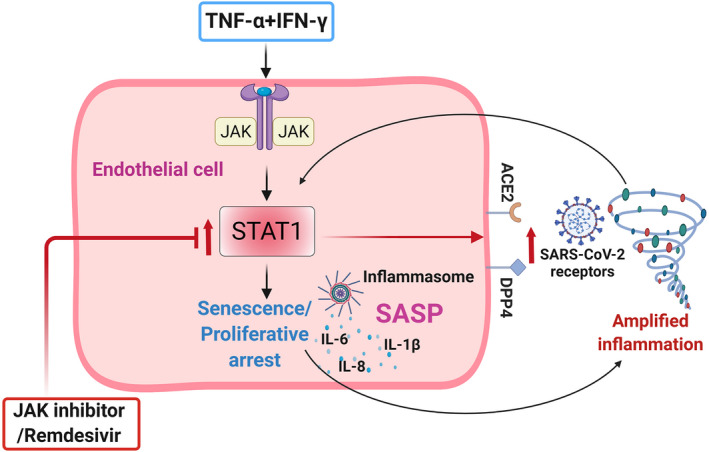
Older age and underlying conditions such as diabetes/obesity or immunosuppression are leading host risk factors for developing severe complications from COVID-19 infection. The pathogenesis of COVID-19-related cytokine storm, tissue damage, and fibrosis may be interconnected with fundamental aging processes, including dysregulated immune responses and cellular senescence. Here, we examined effects of key cytokines linked to cellular senescence on expression of SARS-CoV-2 viral entry receptors. We found exposure of human umbilical vein endothelial cells (HUVECs) to the inflammatory cytokines, TNF-α + IFN-γ or a cocktail of TNF-α + IFN-γ + IL-6, increased expression of ACE2/DPP4, accentuated the pro-inflammatory senescence-associated secretory phenotype (SASP), and decreased cellular proliferative capacity, consistent with progression towards a cellular senescence-like state. IL-6 by itself failed to induce substantial effects on viral entry receptors or SASP-related genes, while synergy between TNF-α and IFN-γ initiated a positive feedback loop via hyper-activation of the JAK/STAT1 pathway, causing SASP amplification. Breaking the interactive loop between senescence and cytokine secretion with JAK inhibitor ruxolitinib or antiviral drug remdesivir prevented hyper-inflammation, normalized SARS-CoV-2 entry receptor expression, and restored HUVECs proliferative capacity. This loop appears to underlie cytokine-mediated viral entry receptor activation and links with senescence and hyper-inflammation.
Keywords: ACE2; COVID-19; DPP4; JAK-STAT; SARS-COV-2 receptor; cytokines; inflammation; senescence.
© 2022 The Authors. Aging Cell published by Anatomical Society and John Wiley & Sons Ltd.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
References
-
- Acosta, J. C. , O'Loghlen, A. , Banito, A. , Guijarro, M. V. , Augert, A. , Raguz, S. , Fumagalli, M. , da Costa, M. , Brown, C. , Popov, N. , Takatsu, Y. , Melamed, J. , d'Adda di Fagagna, F. , Bernard, D. , Hernando, E. , & Gil, J. (2008). Chemokine signaling via the CXCR2 receptor reinforces senescence. Cell, 133(6), 1006–1018. 10.1016/j.cell.2008.03.038 - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
