

Genetic Diversity in Nitrogen Fertiliser Responses and N Gas Emission in Modern Wheat
- PMID: 35646002
- PMCID: PMC9137425
- DOI: 10.3389/fpls.2022.816475
Genetic Diversity in Nitrogen Fertiliser Responses and N Gas Emission in Modern Wheat
Abstract
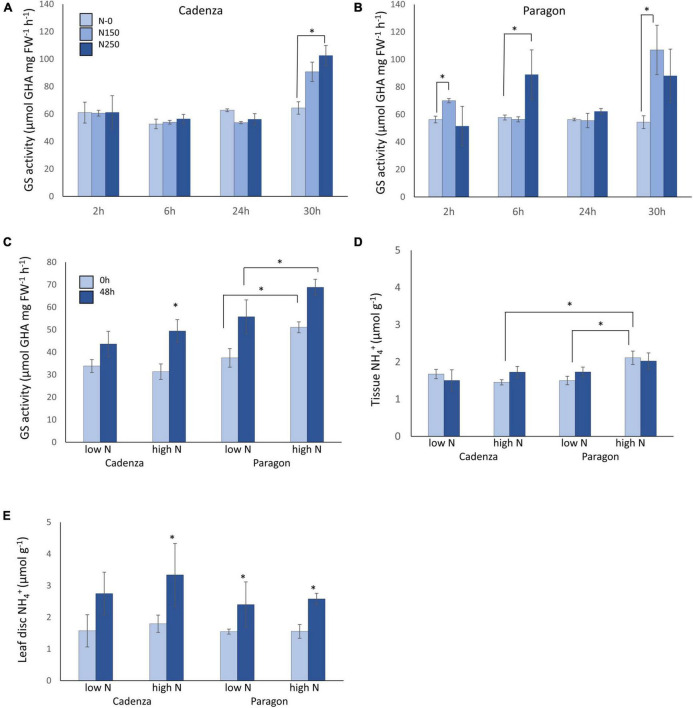
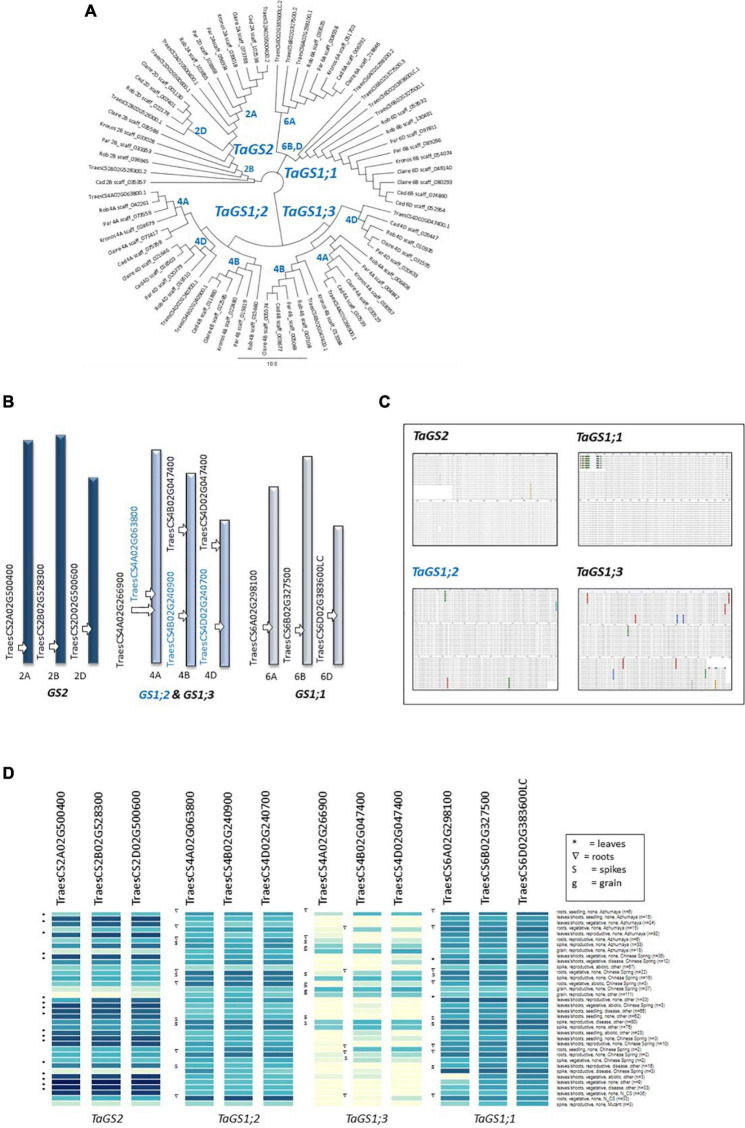
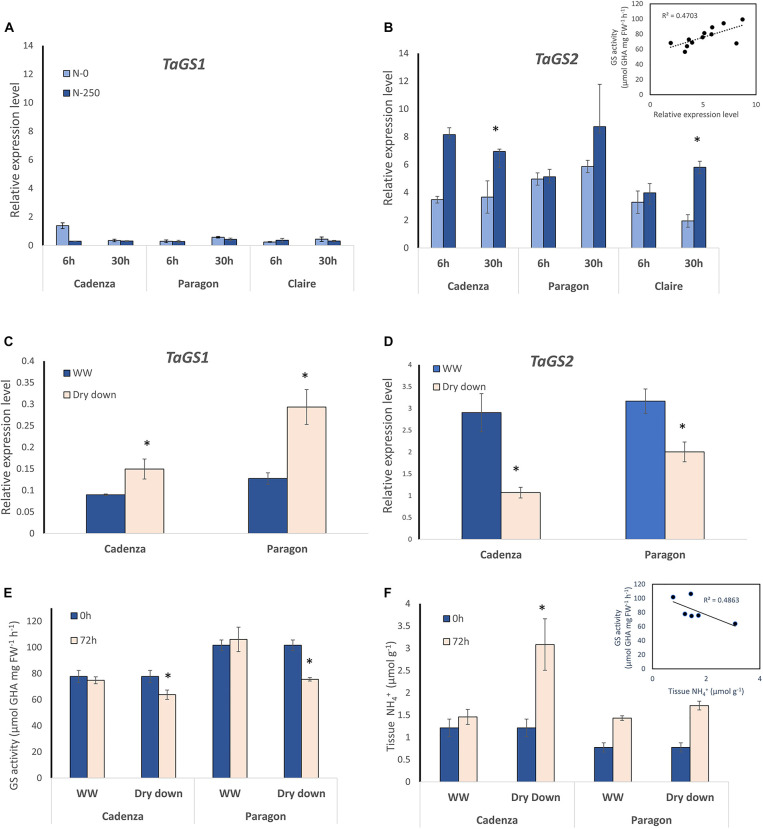
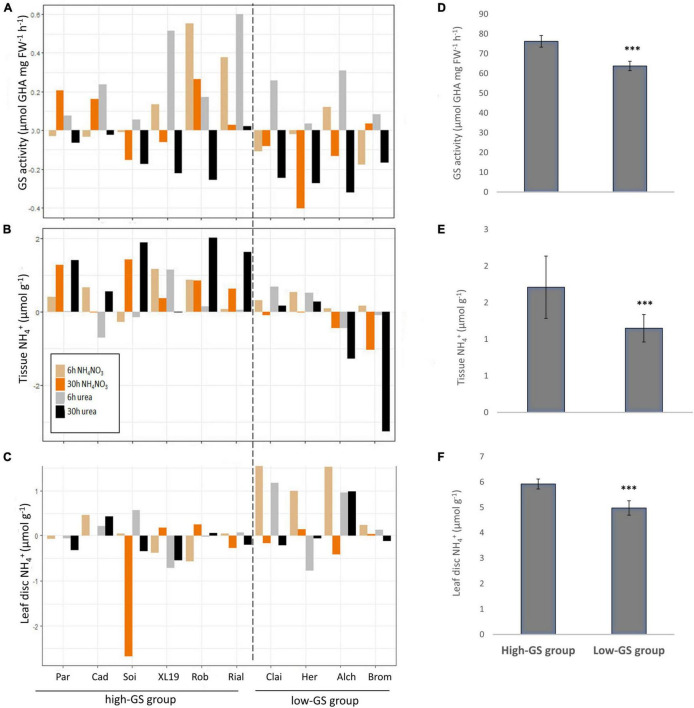
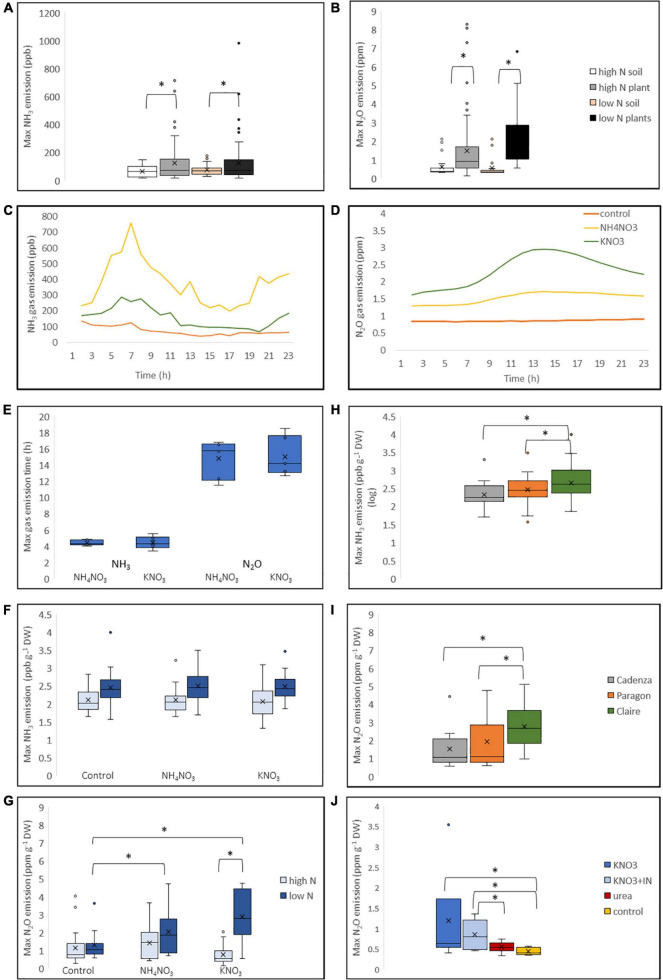
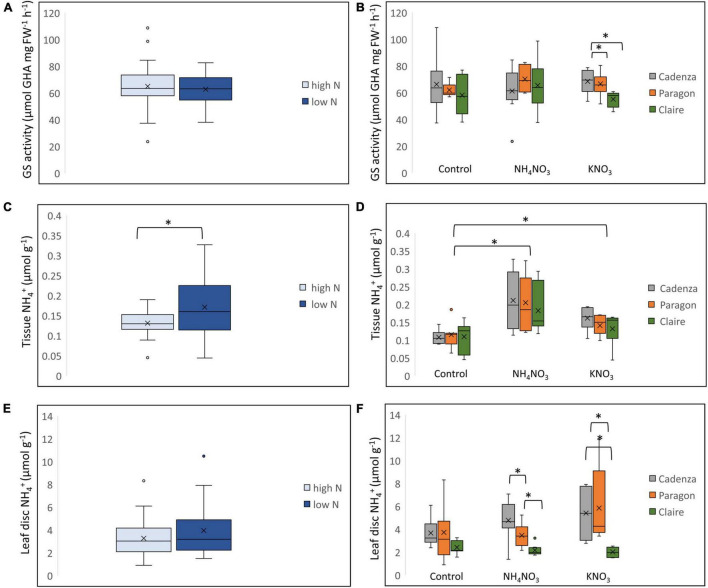
Crops assimilate nitrogen (N) as ammonium via the glutamine synthetase/glutamate synthase (GS/GOGAT) pathway which is of central importance for N uptake and potentially represents a bottle neck for N fertiliser-use efficiency. The aim of this study was to assess whether genetic diversity for N-assimilation capacity exists in wheat and could be exploited for breeding. Wheat plants rapidly, within 6 h, responded to N application with an increase in GS activity. This was not accompanied by an increase in GS gene transcript abundance and a comparison of GS1 and GS2 protein models revealed a high degree of sequence conservation. N responsiveness amongst ten wheat varieties was assessed by measuring GS enzyme activity, leaf tissue ammonium, and by a leaf-disc assay as a proxy for apoplastic ammonia. Based on these data, a high-GS group showing an overall positive response to N could be distinguished from an inefficient, low-GS group. Subsequent gas emission measurements confirmed plant ammonia emission in response to N application and also revealed emission of N2O when N was provided as nitrate, which is in agreement with our current understanding that N2O is a by-product of nitrate reduction. Taken together, the data suggest that there is scope for improving N assimilation capacity in wheat and that further investigations into the regulation and role of GS-GOGAT in NH3 emission is justified. Likewise, emission of the climate gas N2O needs to be reduced, and future research should focus on assessing the nitrate reductase pathway in wheat and explore fertiliser management options.
Keywords: N2O greenhouse gas; Triticum aestivum L; drought; genetic diversity; glutamine synthetase; nitrogen fertiliser.
Copyright © 2022 Oszvald, Hassall, Hughes, Torres-Ballesteros, Clark, Riche and Heuer.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
References
LinkOut - more resources
Full Text Sources