

Corneal stromal repair and regeneration
- PMID: 35649962
- PMCID: PMC11926992
- DOI: 10.1016/j.preteyeres.2022.101090
Corneal stromal repair and regeneration
Abstract
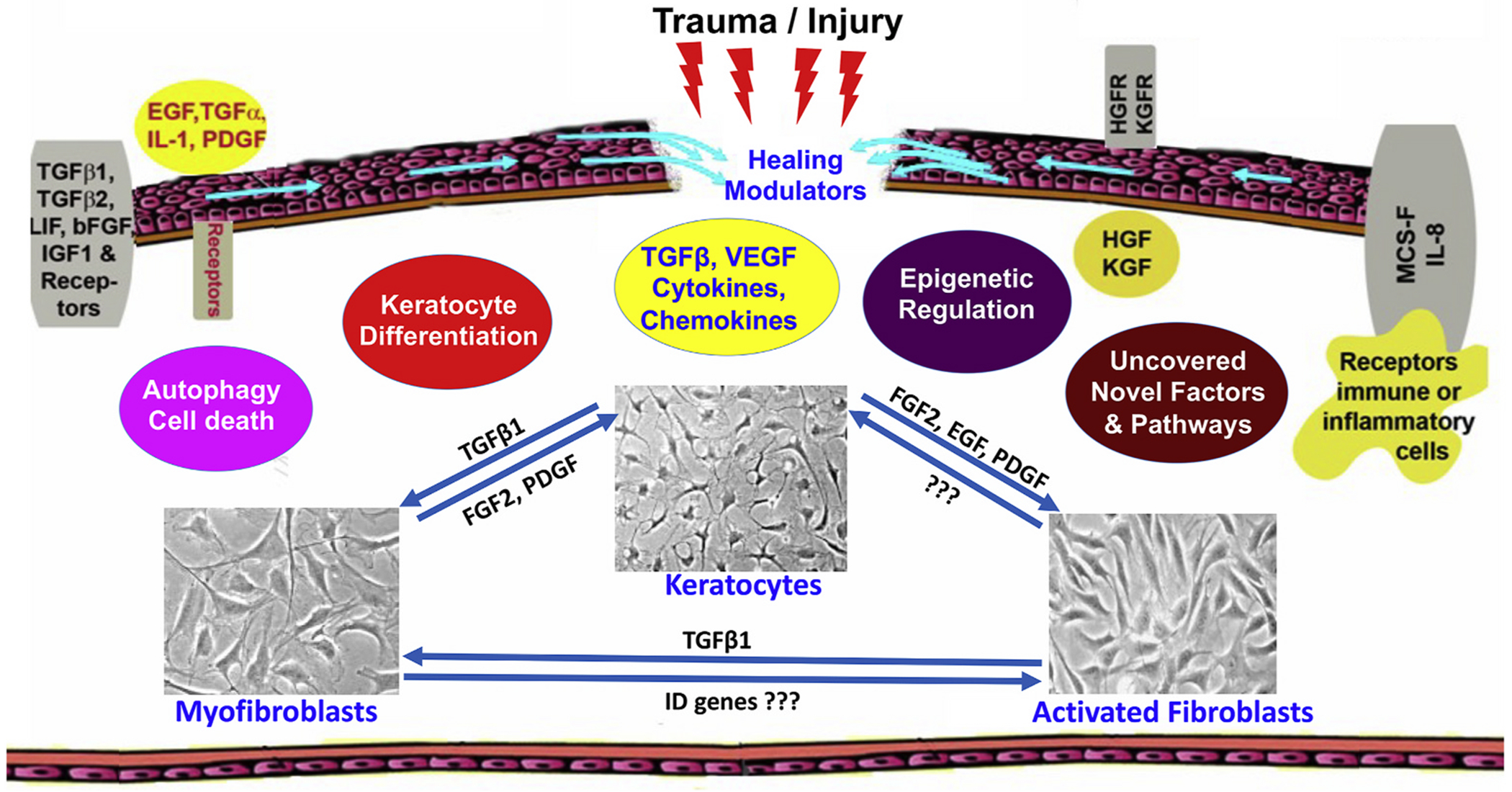
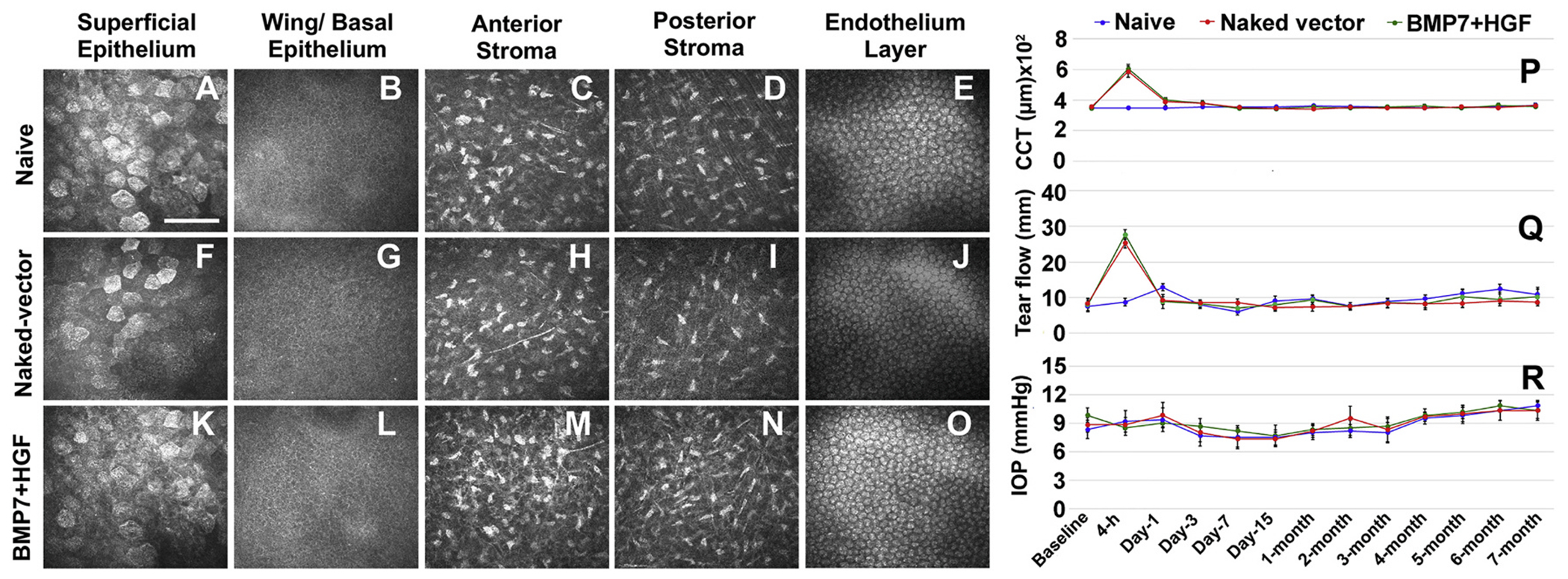
The cornea is a specialized, transparent, avascular, immune-privileged, and heavily innervated tissue that affords 2/3rd of refraction to the eye. Ocular injuries, infections, and genetic factors affect corneal function and cause vision impairment. Presently, a variety of laser/non-laser surgeries, immunosuppressants, and/or corneal transplants are predominantly used to revive sight in human patients. The development of novel, precision-guided, and tissue-targeted non-surgical therapies promoting corneal repair and regeneration based on mechanistic understanding is of paramount importance to reduce the impact of global blindness. Research over the past decade revealed that modulation of pathological signaling pathways and factors by a variety of therapeutic delivery methods effectively treats corneal disorders including corneal scar/haze, inflammation, and angiogenesis in various pre-clinical animal models and are primed for human translation. This review discusses recent advances in the areas of corneal repair, restoration, and regeneration. Herein, we provide an overview of evolving approaches and therapeutic modalities that have shown great promise in reviving corneal transparency and function through the use of small drug molecules, gene therapy, nanomedicine, stem cells, trophic factors, exosomes, stromal equivalents, bioengineered stromal scaffolds, tissue adhesives, and 3D bioprinting.
Keywords: Cornea; Corneal gene therapy; Corneal wound healing; Emerging therapies; Keratocytes; Stromal regeneration; Stromal remodeling.
Copyright © 2022. Published by Elsevier Ltd.
Conflict of interest statement
Declaration of competing interest None.
Figures





















References
-
- Agarwal S, Srinivasan B, Gupta R, Iyer G, 2020. Allogenic simple limbal epithelial transplantation versus amniotic membrane grafting in the early management of severe-grade ocular chemical injuries-A retrospective comparative study. Am. J. Ophthalmol 217, 297–304. - PubMed
-
- Ahmed F, House RJ, Feldman BH, 2015. Corneal abrasions and corneal foreign bodies. Prim Care 42, 363–375. - PubMed
-
- Alio Del Barrio JL, Arnalich-Montiel F, De Miguel MP, El Zarif M, Alio JL, 2021a. Corneal stroma regeneration: preclinical studies. Exp. Eye Res 202, 108314. - PubMed
-
- Alio Del Barrio JL, Bhogal M, Ang M, Ziaei M, Robbie S, Montesel A, Gore DM, Mehta JS, Alio JL, 2021b. Corneal transplantation after failed grafts: options and outcomes. Surv. Ophthalmol 66, 20–40. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
