

De novo biosynthesis of rubusoside and rebaudiosides in engineered yeasts
- PMID: 35650215
- PMCID: PMC9160076
- DOI: 10.1038/s41467-022-30826-2
De novo biosynthesis of rubusoside and rebaudiosides in engineered yeasts
Abstract
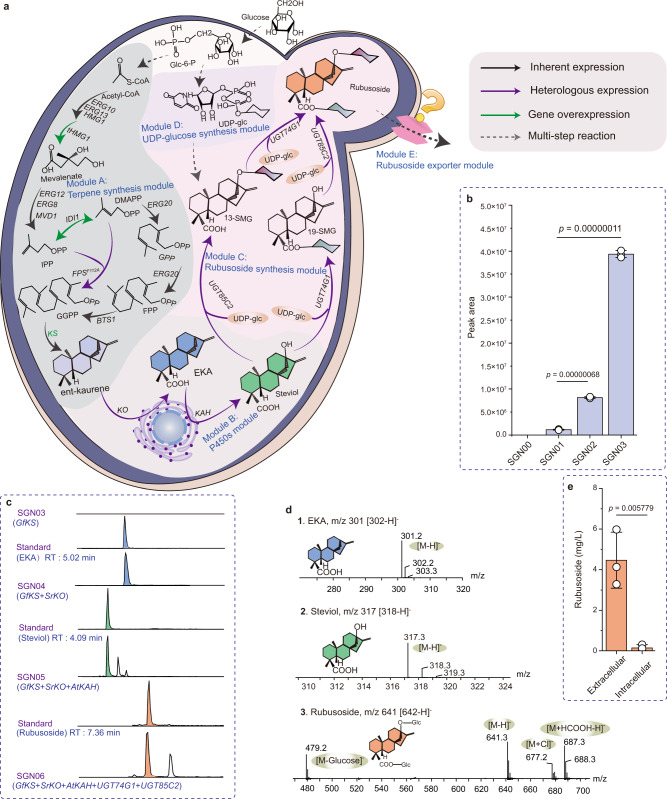
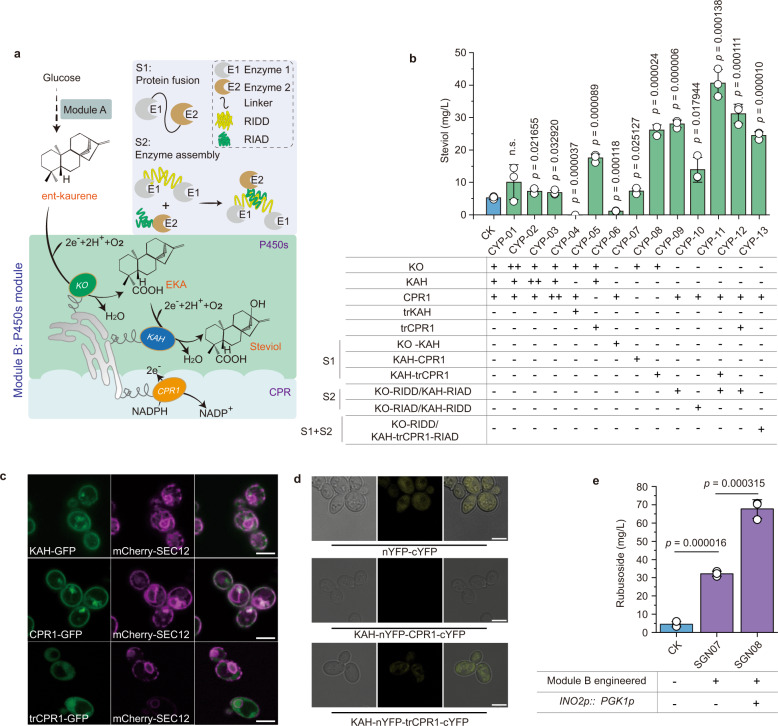
High-sugar diet causes health problems, many of which can be addressed with the use of sugar substitutes. Rubusoside and rebaudiosides are interesting molecules, considered the next generation of sugar substitutes due to their low-calorie, superior sweetness and organoleptic properties. However, their low abundance in nature makes the traditional plant extraction process neither economical nor environmental-friendly. Here we engineer baker's yeast Saccharomyces cerevisiae as a chassis for the de novo production of rubusoside and rebaudiosides. In this process, we identify multiple issues that limit the production, including rate-liming steps, product stress on cellular fitness and unbalanced metabolic networks. We carry out a systematic engineering strategy to solve these issues, which produces rubusoside and rebaudiosides at titers of 1368.6 mg/L and 132.7 mg/L, respectively. The rubusoside chassis strain here constructed paves the way towards a sustainable, large-scale fermentation-based manufacturing of diverse rebaudiosides.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures



Similar articles
-
Enzymatic Monoglucosylation of Rubusoside and the Structure-Sweetness/Taste Relationship of Monoglucosyl Derivatives.J Agric Food Chem. 2020 Aug 12;68(32):8702-8709. doi: 10.1021/acs.jafc.0c03236. Epub 2020 Jul 30. J Agric Food Chem. 2020. PMID: 32686405
-
Selective production of rubusoside from stevioside by using the sophorose activity of β-glucosidase from Streptomyces sp. GXT6.Appl Microbiol Biotechnol. 2015 Nov;99(22):9663-74. doi: 10.1007/s00253-015-6802-z. Epub 2015 Jul 22. Appl Microbiol Biotechnol. 2015. PMID: 26198882
-
Production of rubusoside from stevioside by using a thermostable lactase from Thermus thermophilus and solubility enhancement of liquiritin and teniposide.Enzyme Microb Technol. 2014 Oct;64-65:38-43. doi: 10.1016/j.enzmictec.2014.07.001. Epub 2014 Jul 11. Enzyme Microb Technol. 2014. PMID: 25152415
-
Harnessing the yeast Saccharomyces cerevisiae for the production of fungal secondary metabolites.Essays Biochem. 2021 Jul 26;65(2):277-291. doi: 10.1042/EBC20200137. Essays Biochem. 2021. PMID: 34061167 Free PMC article. Review.
-
Metabolic engineering of Saccharomyces cerevisiae for lactose/whey fermentation.Bioeng Bugs. 2010 May-Jun;1(3):164-71. doi: 10.4161/bbug.1.3.10619. Epub 2009 Nov 13. Bioeng Bugs. 2010. PMID: 21326922 Free PMC article. Review.
Cited by
-
Biosynthesis and metabolic engineering of natural sweeteners.AMB Express. 2025 Mar 18;15(1):50. doi: 10.1186/s13568-025-01864-y. AMB Express. 2025. PMID: 40100508 Free PMC article. Review.
-
Enhanced Production of Rebaudioside D and Rebaudioside M through V155T Substitution in the Glycosyltransferase UGT91D2 from Stevia rebaudiana.J Agric Food Chem. 2025 Jan 22;73(3):2019-2032. doi: 10.1021/acs.jafc.4c09392. Epub 2025 Jan 9. J Agric Food Chem. 2025. PMID: 39783863 Free PMC article.
-
Sustainable biosynthesis of valuable diterpenes in microbes.Eng Microbiol. 2022 Nov 10;3(1):100058. doi: 10.1016/j.engmic.2022.100058. eCollection 2023 Mar. Eng Microbiol. 2022. PMID: 39628524 Free PMC article. Review.
-
Yeast metabolism adaptation for efficient terpenoids synthesis via isopentenol utilization.Nat Commun. 2024 Nov 13;15(1):9844. doi: 10.1038/s41467-024-54298-8. Nat Commun. 2024. PMID: 39537637 Free PMC article.
-
Rational Design for the Complete Synthesis of Stevioside in Saccharomyces cerevisiae.Microorganisms. 2024 May 31;12(6):1125. doi: 10.3390/microorganisms12061125. Microorganisms. 2024. PMID: 38930507 Free PMC article.
References
-
- World Health Organization. Global action plan for the prevention and control of noncommunicable diseases 2013–2020. (World Health Organization, 2013).
-
- Zhang J, Chou G, Liu Z, Liu M. Employing rubusoside to improve the solubility and permeability of antitumor compound betulonic acid. Nanomedicine. 2016;11:2829–2844. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
