

An oxygen-sensing mechanism for angiosperm adaptation to altitude
- PMID: 35650430
- PMCID: PMC9200633
- DOI: 10.1038/s41586-022-04740-y
An oxygen-sensing mechanism for angiosperm adaptation to altitude
Abstract
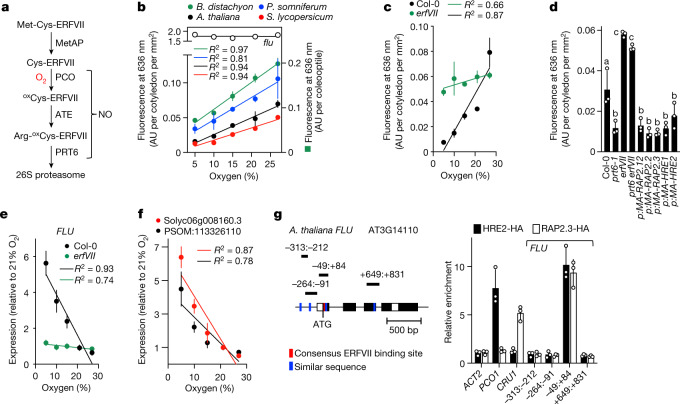
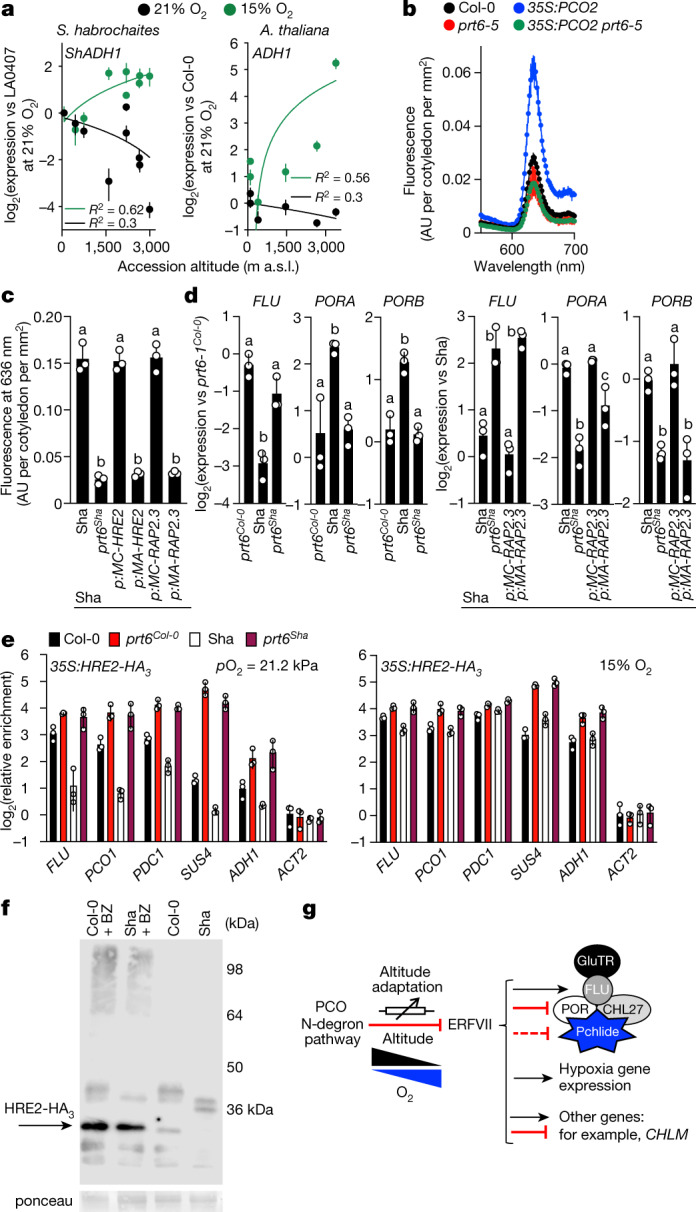
Flowering plants (angiosperms) can grow at extreme altitudes, and have been observed growing as high as 6,400 metres above sea level1,2; however, the molecular mechanisms that enable plant adaptation specifically to altitude are unknown. One distinguishing feature of increasing altitude is a reduction in the partial pressure of oxygen (pO2). Here we investigated the relationship between altitude and oxygen sensing in relation to chlorophyll biosynthesis-which requires molecular oxygen3-and hypoxia-related gene expression. We show that in etiolated seedlings of angiosperm species, steady-state levels of the phototoxic chlorophyll precursor protochlorophyllide are influenced by sensing of atmospheric oxygen concentration. In Arabidopsis thaliana, this is mediated by the PLANT CYSTEINE OXIDASE (PCO) N-degron pathway substrates GROUP VII ETHYLENE RESPONSE FACTOR transcription factors (ERFVIIs). ERFVIIs positively regulate expression of FLUORESCENT IN BLUE LIGHT (FLU), which represses the first committed step of chlorophyll biosynthesis, forming an inactivation complex with tetrapyrrole synthesis enzymes that are negatively regulated by ERFVIIs, thereby suppressing protochlorophyllide. In natural populations representing diverse angiosperm clades, we find oxygen-dependent altitudinal clines for steady-state levels of protochlorophyllide, expression of inactivation complex components and hypoxia-related genes. Finally, A. thaliana accessions from contrasting altitudes display altitude-dependent ERFVII activity and accumulation. We thus identify a mechanism for genetic adaptation to absolute altitude through alteration of the sensitivity of the oxygen-sensing system.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures











Comment in
-
High on oxygen.Nat Plants. 2022 Jul;8(7):731-732. doi: 10.1038/s41477-022-01196-w. Nat Plants. 2022. PMID: 35773418 No abstract available.
References
-
- Dentant C. The highest vascular plants on Earth. Alp. Bot. 2018;128:97–106. doi: 10.1007/s00035-018-0208-3. - DOI
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
