

The Water to Land Transition Submerged: Multifunctional Design of Pectoral Fins for Use in Swimming and in Association with Underwater Substrate
- PMID: 35652788
- PMCID: PMC9617210
- DOI: 10.1093/icb/icac061
The Water to Land Transition Submerged: Multifunctional Design of Pectoral Fins for Use in Swimming and in Association with Underwater Substrate
Abstract
Fins of fishes provide many examples of structures that are beautifully designed to power and control movement in water; however, some species also use their fins for substrate-associated behaviors where interactions with solid surfaces are key. Here, we examine how the pectoral fins of ray-finned fish with these multifunctional behavioral demands, in water and on solid surfaces, are structured and function. We subdivide fins used in swimming and substrate contact into two general morphological categories, regionalized vs. generalized fins. Regionalized fins have ventral rays that are free from connecting membrane or in which that membrane is reduced. Dorsally they maintain a more typical membranous fin. While all pectoral fins vary somewhat in their morphology from leading to trailing edge, generalized fins do not have the substantial membrane loss between rays that is seen in regionalized fins and the distal edge anatomy changes gradually along its margin. We add a new case study in regionalized fins with the dwarf hawkfish (Cirrhitichthys falco). Hawkfishes are most often found perching and moving on structures in their environments. During perching, the free ventral rays are in contact with the substrate and splayed. We found that unlike other fish with regionalized pectoral fins, hawkfish maintain use of the dorsal membranous region of its pectoral fin for rhythmic swimming. We found that typically hawkfish bend their ventral free rays under, toward the medial hemitrichs or hold them straight during substrate-associated postures. This appears also to be the case for the ventral free rays of other species with regionalized fins. Generalized fin use for substrate contact was reviewed in round gobies (Neogobius melanostomus). In addition, although their lobe fins are not representative of ray-finned fish anatomy, we explored fin contact on submerged substrates in the Senegal bichir (Polypterus senegalus), which has a generalized distal fin (no free fin rays or distinct membrane regions). Both groups use their pectoral fins for swimming. During substrate-based postures, unlike hawkfish, their distal rays generally bend outward toward the lateral hemitrichs and a large swath of the fin membrane can contact the surface. The alternative demands on multifunctional fins suggest specialization of the mechanosensory system. We review mechanosensation related to fin movement and surface contact. These alternative regionalized and generalized strategies for serving aquatic and substrate-based functions underwater provide opportunities to further investigate specializations, including sensory structures and systems, that accompany the evolution of substrate-based behaviors in vertebrates.
© The Author(s) 2022. Published by Oxford University Press on behalf of the Society for Integrative and Comparative Biology.
Figures
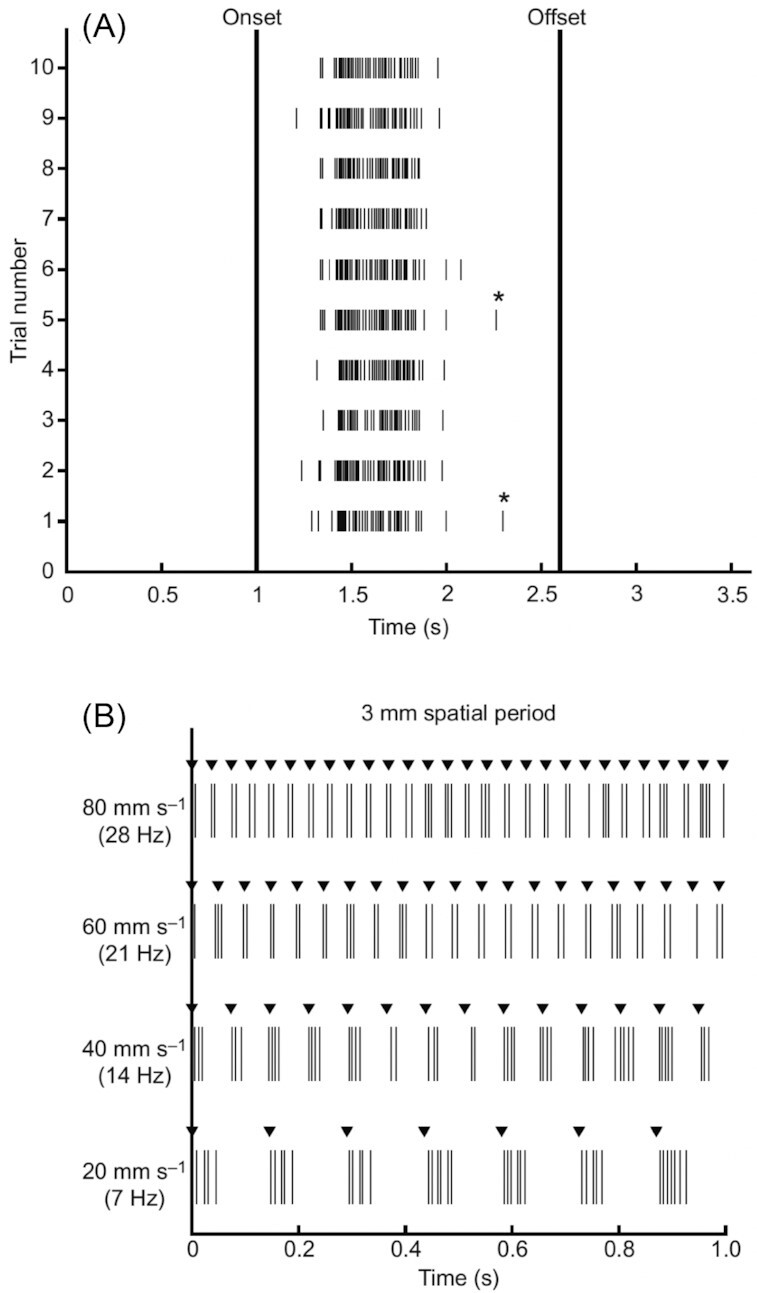
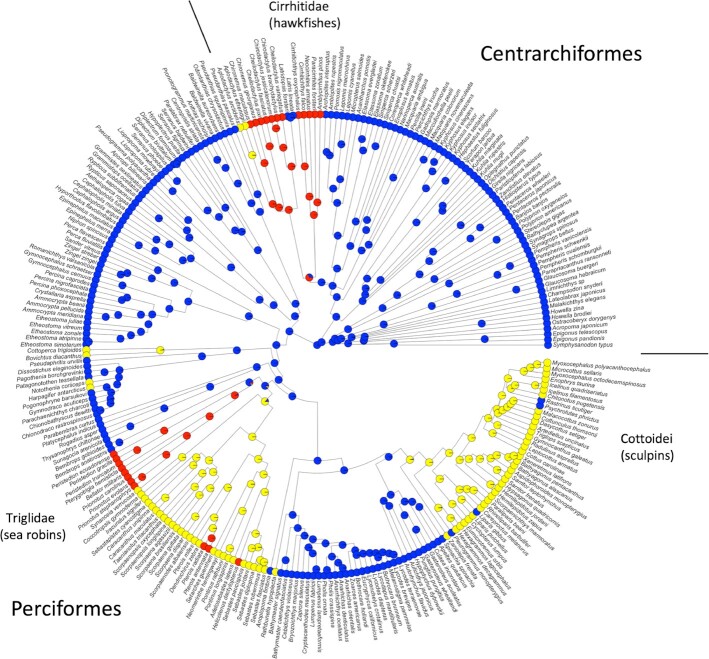
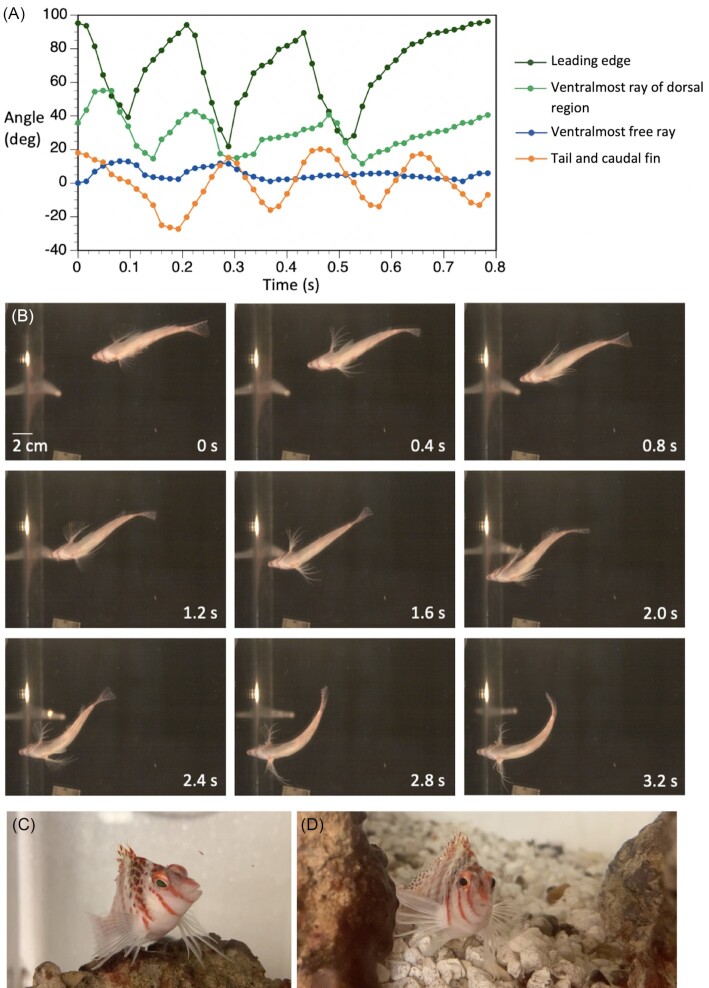

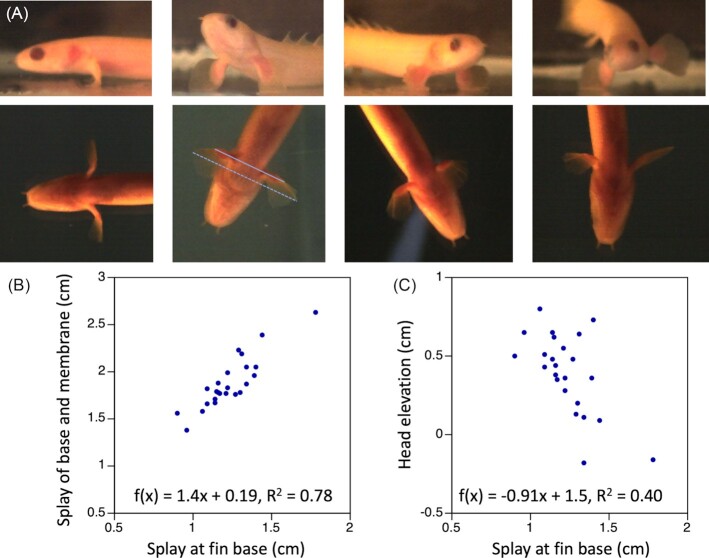
References
-
- Aiello BR, Olsen AM, Mathis CE, Westneat MW, Hale ME. 2020. Pectoral fin kinematics and motor patterns are shaped by fin ray mechanosensation during steady swimming in Scarus quoyi. J Exp Biol. 223:jeb211466. - PubMed
-
- Arreola VI, Westneat MW.. 1996. Mechanics of propulsion by multiple fins: kinematics of aquatic locomotion in the burrfish (Chilomycterus schoepfi). Proc R Soc Lond B. 263:1689–96.
-
- Bardach JE, Case J.. 1965. Sensory capabilities of the modified fins of squirrel hake (Urophycis chuss) and Searobins (Prionotus carolinus and P. evolans). Copeia. 1965:194–206.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
