

The aryl hydrocarbon receptor instructs the immunomodulatory profile of a subset of Clec4a4+ eosinophils unique to the small intestine
- PMID: 35653568
- PMCID: PMC9191779
- DOI: 10.1073/pnas.2204557119
The aryl hydrocarbon receptor instructs the immunomodulatory profile of a subset of Clec4a4+ eosinophils unique to the small intestine
Abstract
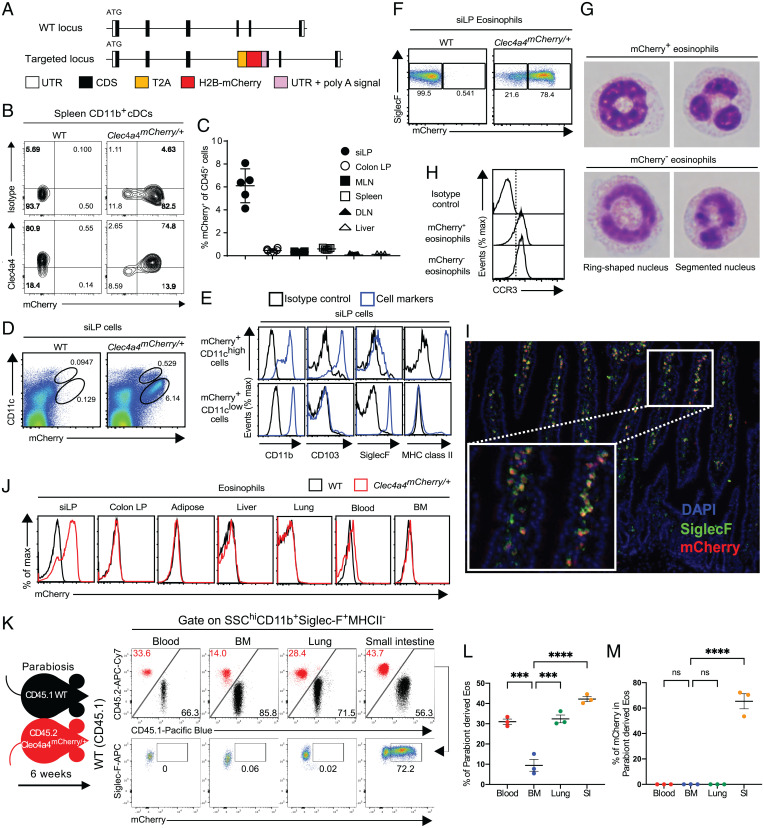
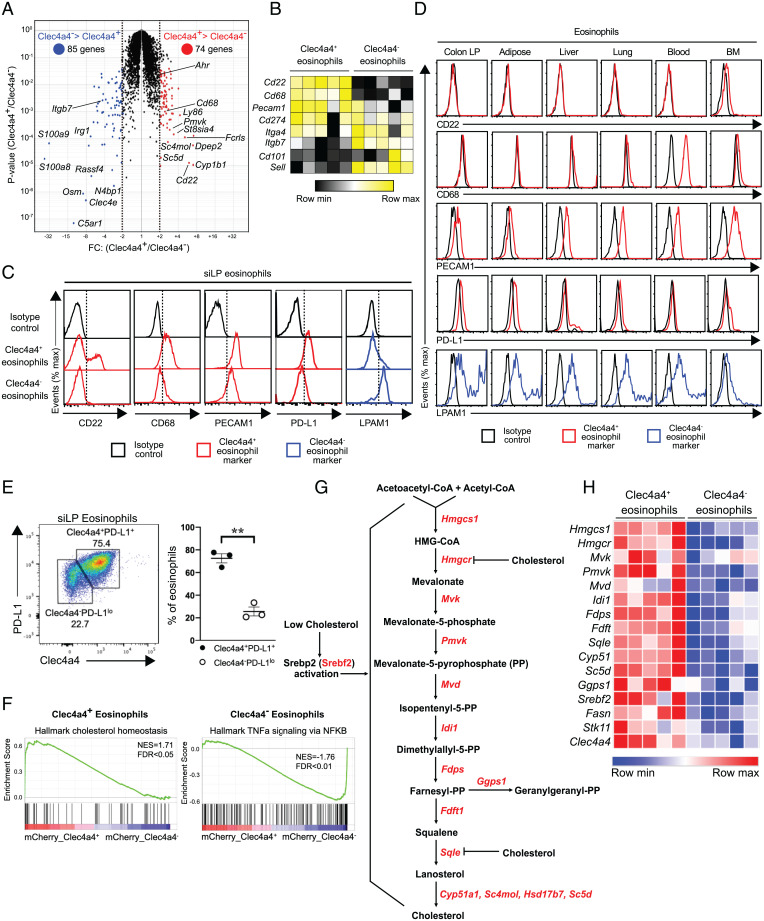
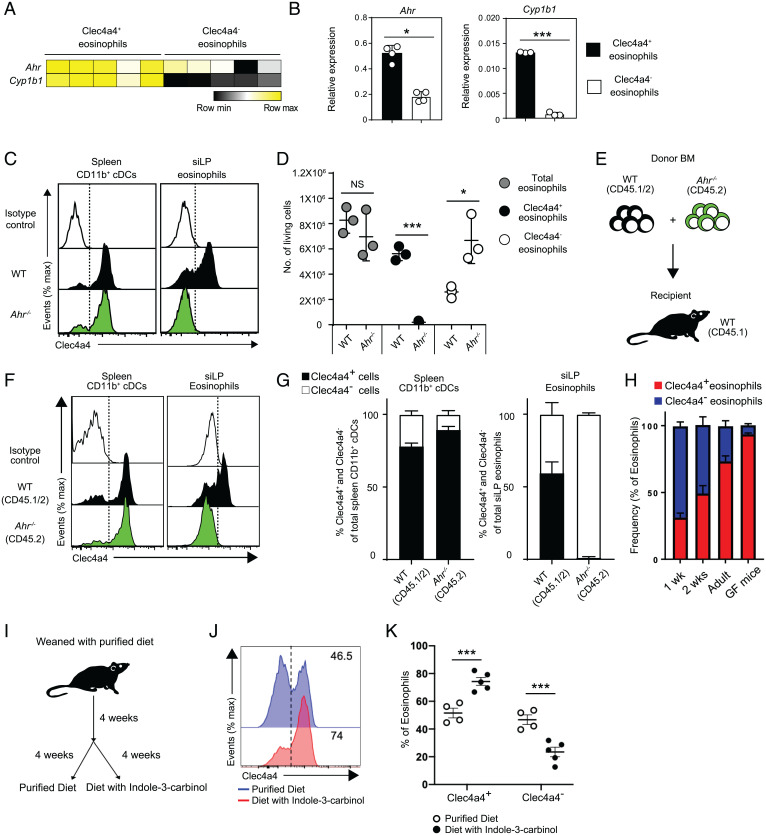
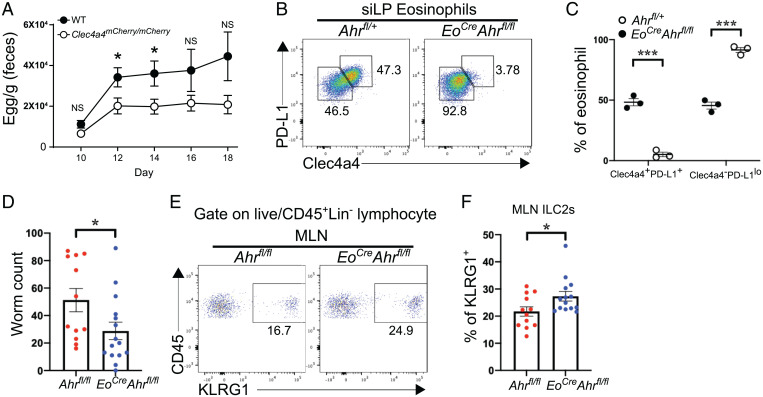
C-type lectin domain family 4, member a4 (Clec4a4) is a C-type lectin inhibitory receptor specific for glycans thought to be exclusively expressed on murine CD8α− conventional dendritic cells. Using newly generated Clec4a4-mCherry knock-in mice, we identify a subset of Clec4a4-expressing eosinophils uniquely localized in the small intestine lamina propria. Clec4a4+ eosinophils evinced an immunomodulatory signature, whereas Clec4a4− eosinophils manifested a proinflammatory profile. Clec4a4+ eosinophils expressed high levels of aryl hydrocarbon receptor (Ahr), which drove the expression of Clec4a4 as well as other immunomodulatory features, such as PD-L1. The abundance of Clec4a4+ eosinophils was dependent on dietary AHR ligands, increased with aging, and declined in inflammatory conditions. Mice lacking AHR in eosinophils expanded innate lymphoid cells of type 2 and cleared Nippostrongylus brasiliensis infection more effectively than did wild-type mice. These results highlight the heterogeneity of eosinophils in response to tissue cues and identify a unique AHR-dependent subset of eosinophils in the small intestine with an immunomodulatory profile.
Keywords: allergy; aryl hydrocarbon receptor; eosinophil; helminth; intestine.
Conflict of interest statement
Competing interest statement: M. Colonna receives research support from Pfizer, Aclaris, Ono, NGM Biopharmaceutical, Oncorus, and VigilNeuro; serves on the scientific advisory boards of Vigil Neuro and NGM Biopharmaceutical; and is consultant for Cell Signaling Technologies. All other authors have nothing to disclose.
Figures
References
-
- Rothenberg M. E., Hogan S. P., The eosinophil. Annu. Rev. Immunol. 24, 147–174 (2006). - PubMed
-
- Gleich G. J., Adolphson C. R., Leiferman K. M., The biology of the eosinophilic leukocyte. Annu. Rev. Med. 44, 85–101 (1993). - PubMed
-
- Hamann K. J., et al. , In vitro killing of microfilariae of Brugia pahangi and Brugia malayi by eosinophil granule proteins. J. Immunol. 144, 3166–3173 (1990). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
