

Complex effects on CaV2.1 channel gating caused by a CACNA1A variant associated with a severe neurodevelopmental disorder
- PMID: 35655070
- PMCID: PMC9163077
- DOI: 10.1038/s41598-022-12789-y
Complex effects on CaV2.1 channel gating caused by a CACNA1A variant associated with a severe neurodevelopmental disorder
Abstract
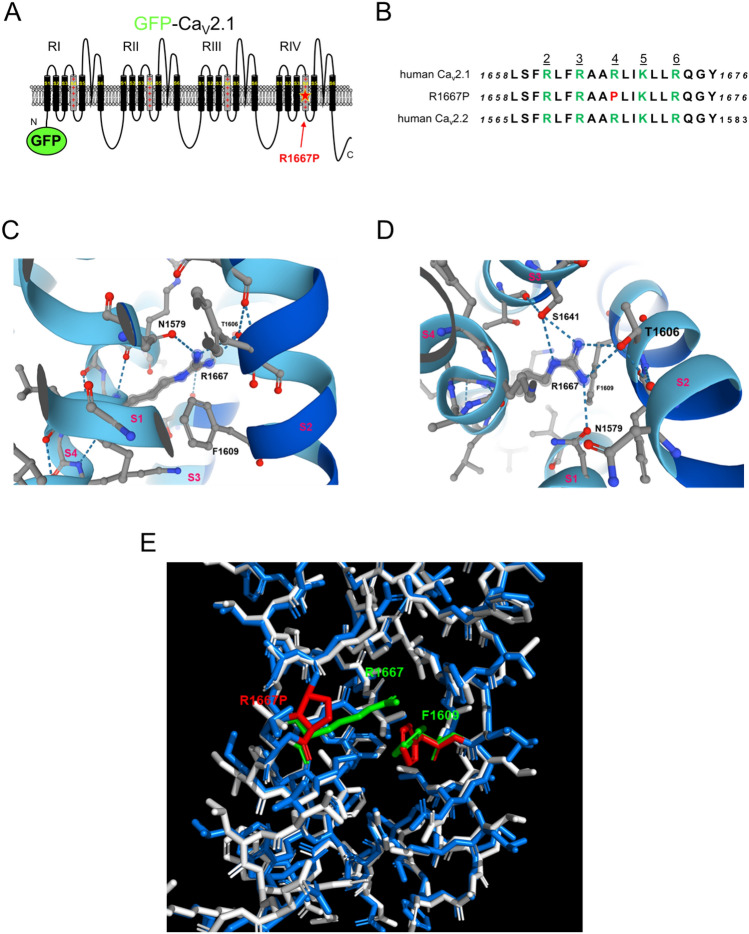
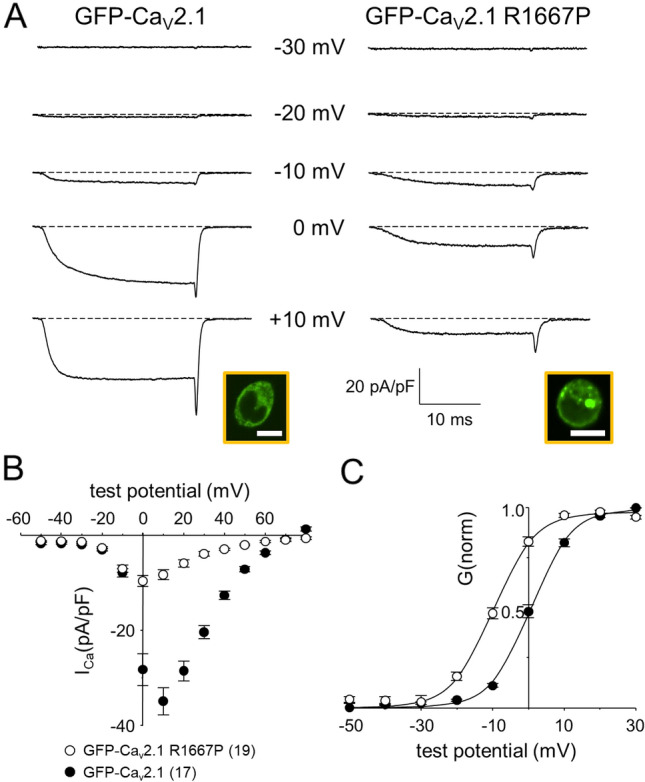
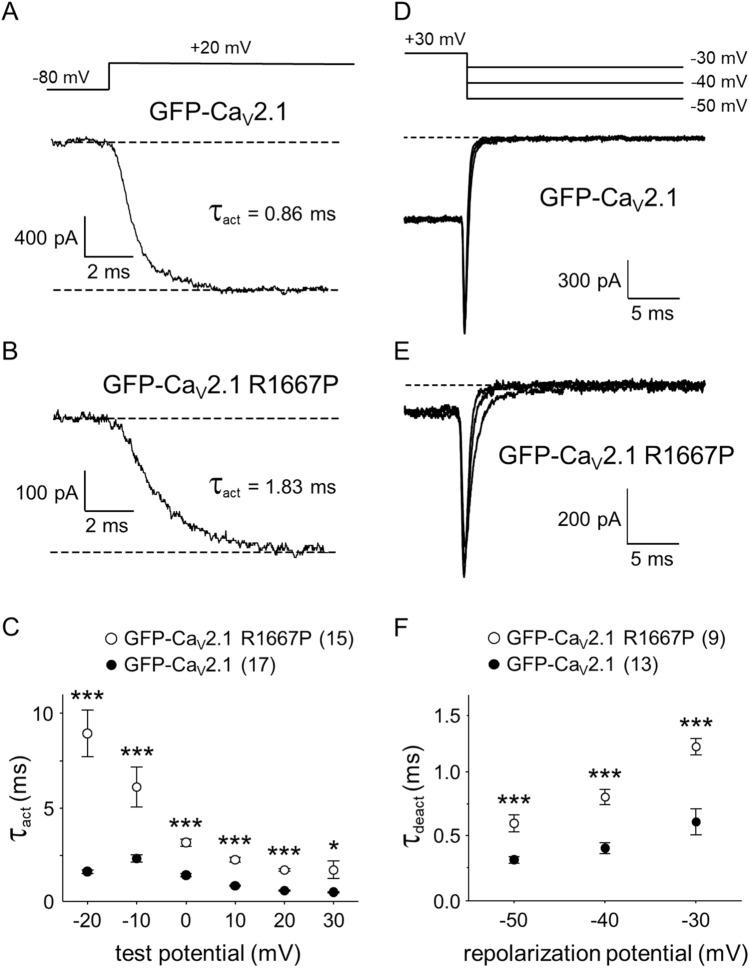
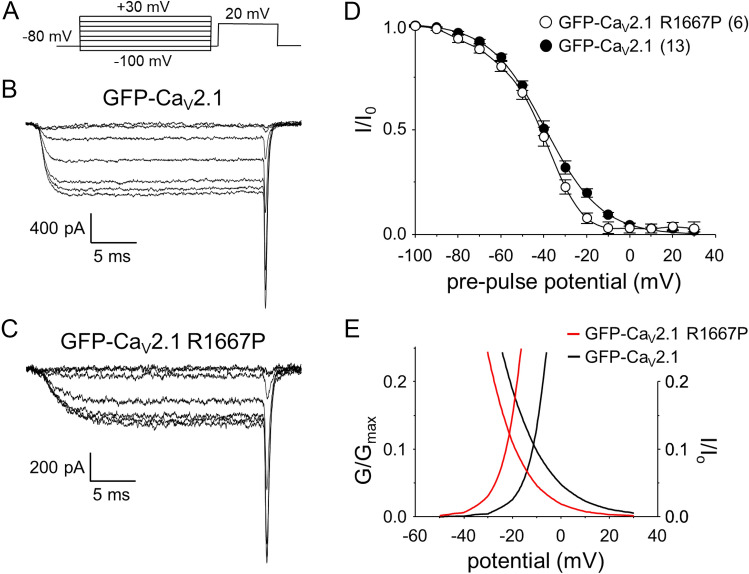
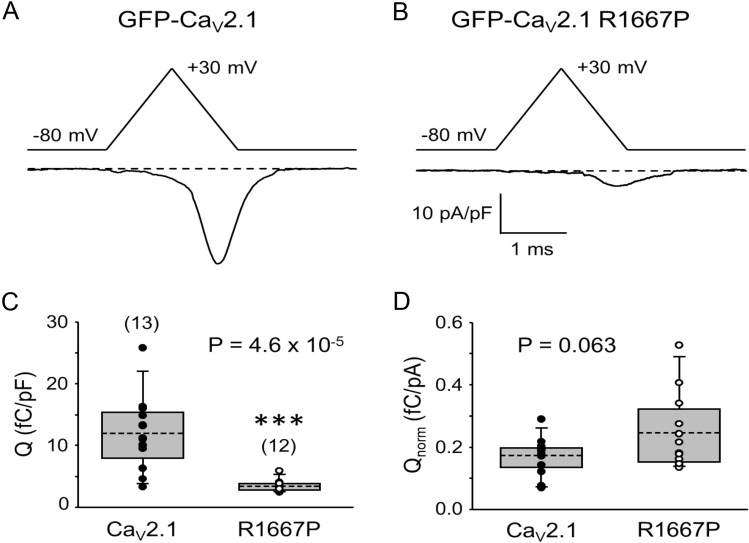
P/Q-type Ca2+ currents mediated by CaV2.1 channels are essential for active neurotransmitter release at neuromuscular junctions and many central synapses. Mutations in CACNA1A, the gene encoding the principal CaV2.1 α1A subunit, cause a broad spectrum of neurological disorders. Typically, gain-of-function (GOF) mutations are associated with migraine and epilepsy while loss-of-function (LOF) mutations are causative for episodic and congenital ataxias. However, a cluster of severe CaV2.1 channelopathies have overlapping presentations which suggests that channel dysfunction in these disorders cannot always be defined bimodally as GOF or LOF. In particular, the R1667P mutation causes focal seizures, generalized hypotonia, dysarthria, congenital ataxia and, in one case, cerebral edema leading ultimately to death. Here, we demonstrate that the R1667P mutation causes both channel GOF (hyperpolarizing voltage-dependence of activation, slowed deactivation) and LOF (slowed activation kinetics) when expressed heterologously in tsA-201 cells. We also observed a substantial reduction in Ca2+ current density in this heterologous system. These changes in channel gating and availability/expression manifested in diminished Ca2+ flux during action potential-like stimuli. However, the integrated Ca2+ fluxes were no different when normalized to tail current amplitude measured upon repolarization from the reversal potential. In summary, our findings indicate a complex functional effect of R1667P and support the idea that pathological missense mutations in CaV2.1 may not represent exclusively GOF or LOF.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
