

Experimental evolution of Bacillus subtilis on Arabidopsis thaliana roots reveals fast adaptation and improved root colonization
- PMID: 35663012
- PMCID: PMC9157203
- DOI: 10.1016/j.isci.2022.104406
Experimental evolution of Bacillus subtilis on Arabidopsis thaliana roots reveals fast adaptation and improved root colonization
Abstract
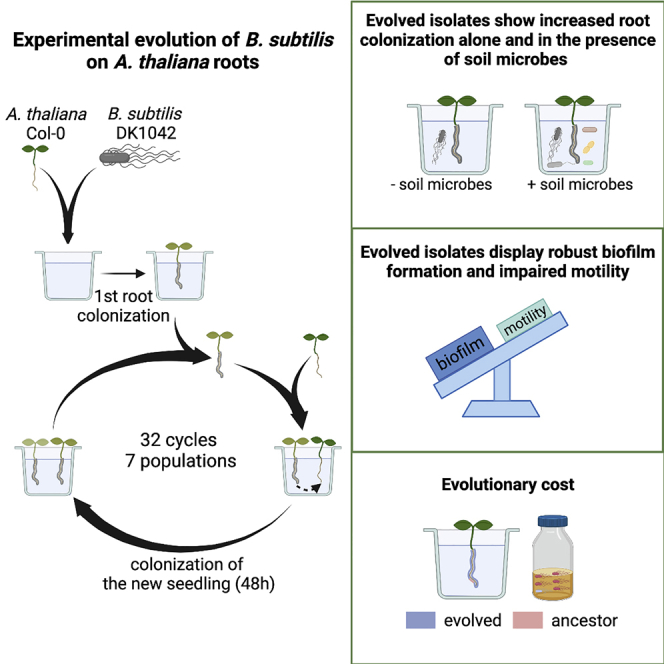
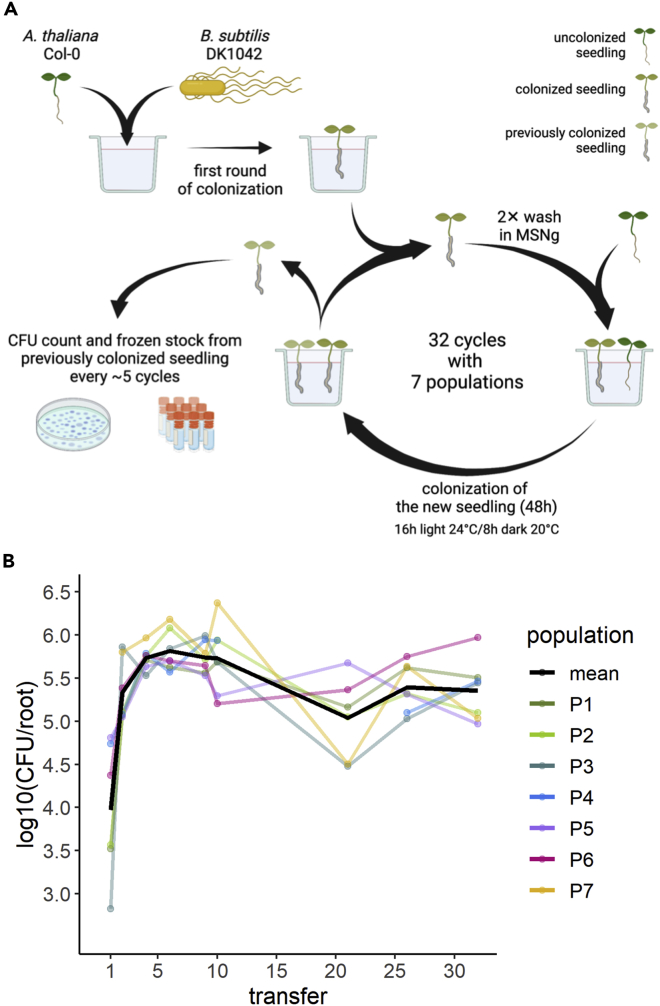
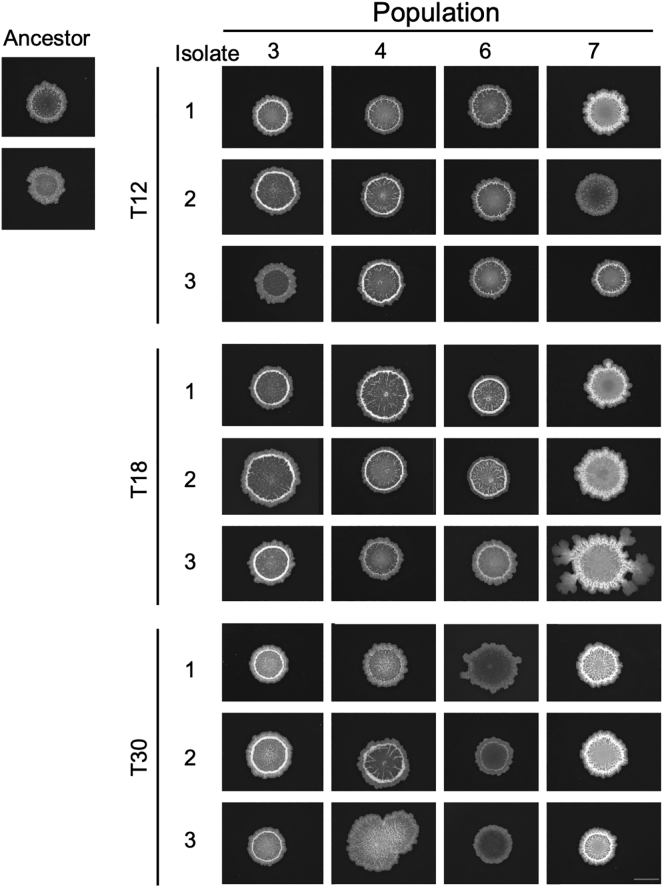
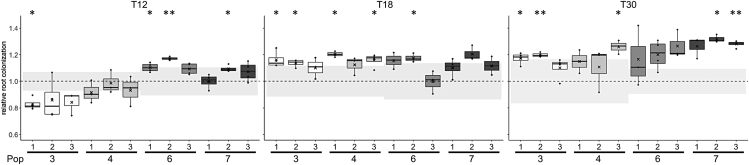
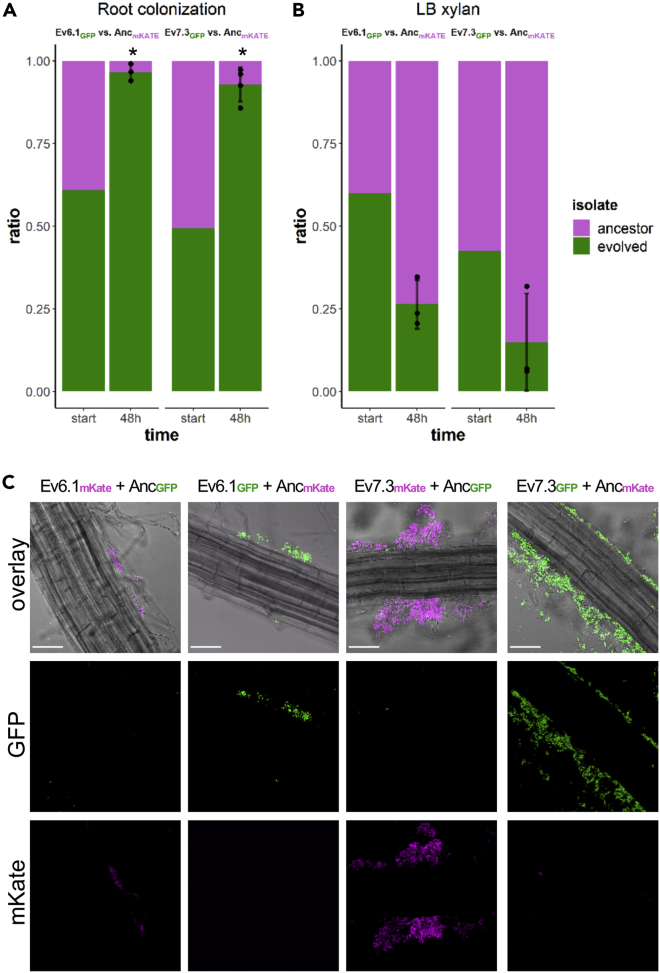
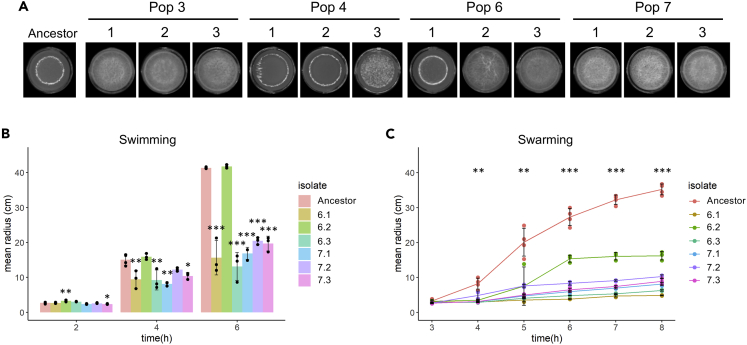
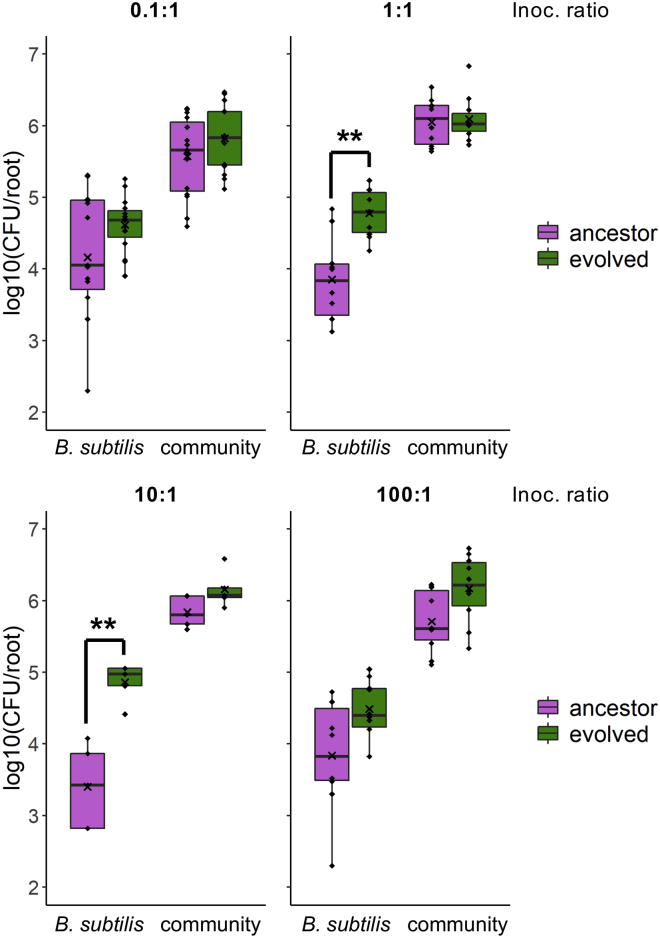
Bacillus subtilis is known to promote plant growth and protect plants against disease. B. subtilis rapidly adapts to Arabidopsis thaliana root colonization, as evidenced by improved root colonizers already after 12 consecutive transfers between seedlings in a hydroponic setup. Re-sequencing of single evolved isolates and endpoint populations revealed mutations in genes related to different bacterial traits, in accordance with evolved isolates displaying increased root colonization associated with robust biofilm formation in response to the plant polysaccharide xylan and impaired motility. Interestingly, evolved isolates suffered a fitness disadvantage in a non-selective environment, demonstrating an evolutionary cost of adaptation to the plant root. Finally, increased root colonization by an evolved isolate was also demonstrated in the presence of resident soil microbes. Our findings highlight how a plant growth-promoting rhizobacterium rapidly adapts to an ecologically relevant environment and reveal evolutionary consequences that are fundamental to consider when evolving strains for biocontrol purposes.
Keywords: ecology; microbiology; plant biology.
© 2022 The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Akram W., Anjum T., Ali B. Searching ISR determinant/s from Bacillus subtilis IAGS174 against Fusarium wilt of tomato. BioControl. 2015;60:271–280. doi: 10.1007/s10526-014-9636-1. - DOI
-
- Allard-Massicotte R., Tessier L., Lécuyer F., Lakshmanan V., Lucier J.F., Garneau D., Caudwell L., Vlamakis H., Bais H.P., Beauregard P.B. Bacillus subtilis early colonization of Arabidopsis thaliana roots involves multiple chemotaxis receptors. MBio. 2016;7:e01664-16. doi: 10.1128/mBio.01664-16. - DOI - PMC - PubMed
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
