

Widespread phages of endosymbionts: Phage WO genomics and the proposed taxonomic classification of Symbioviridae
- PMID: 35666732
- PMCID: PMC9203015
- DOI: 10.1371/journal.pgen.1010227
Widespread phages of endosymbionts: Phage WO genomics and the proposed taxonomic classification of Symbioviridae
Abstract
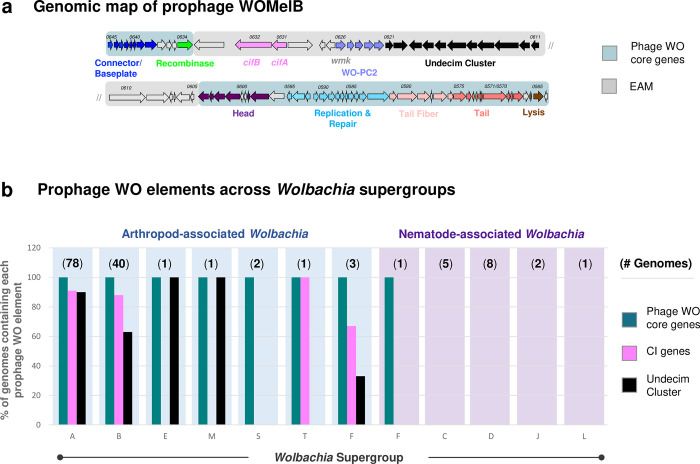
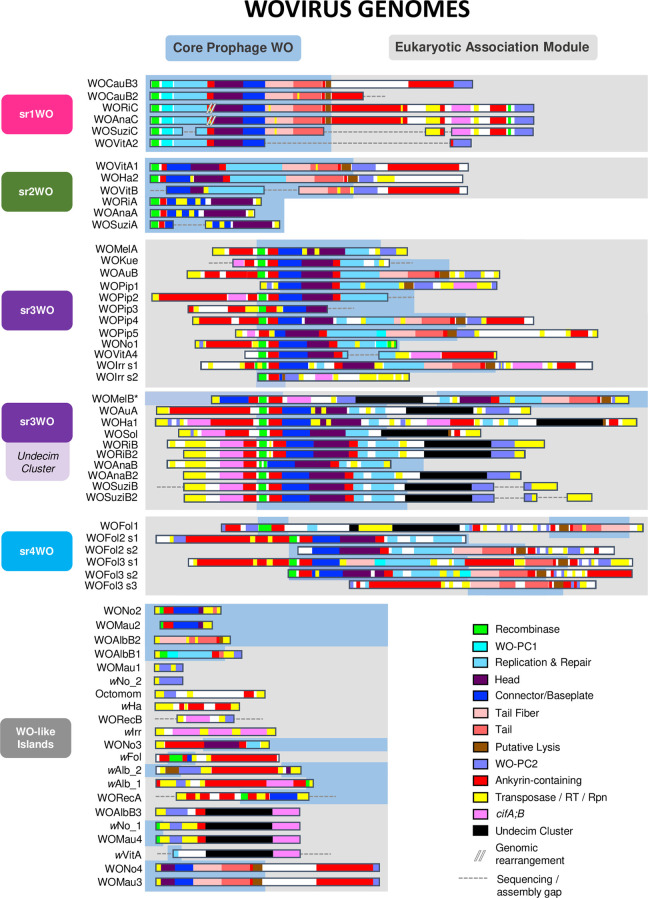
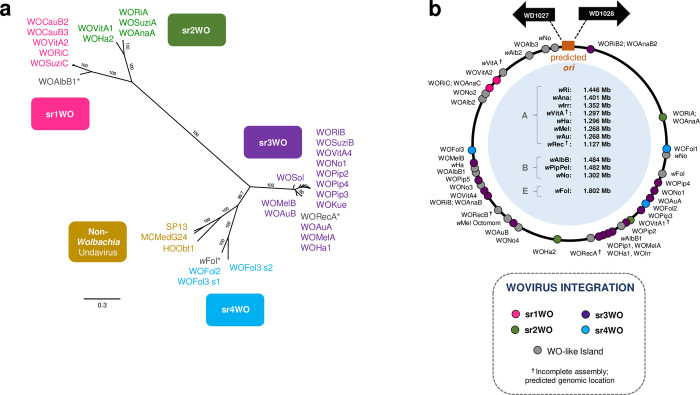
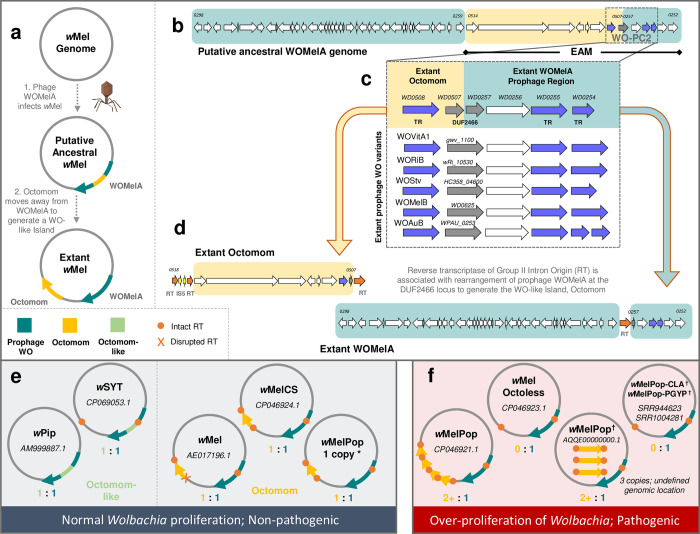
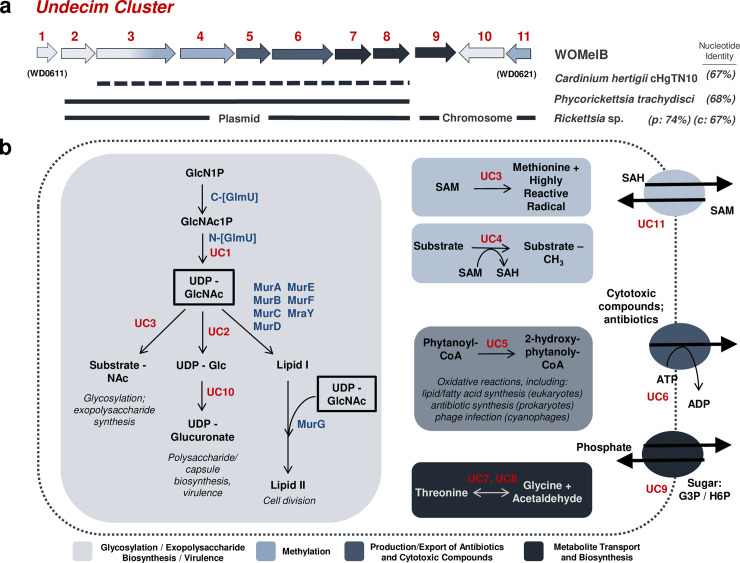
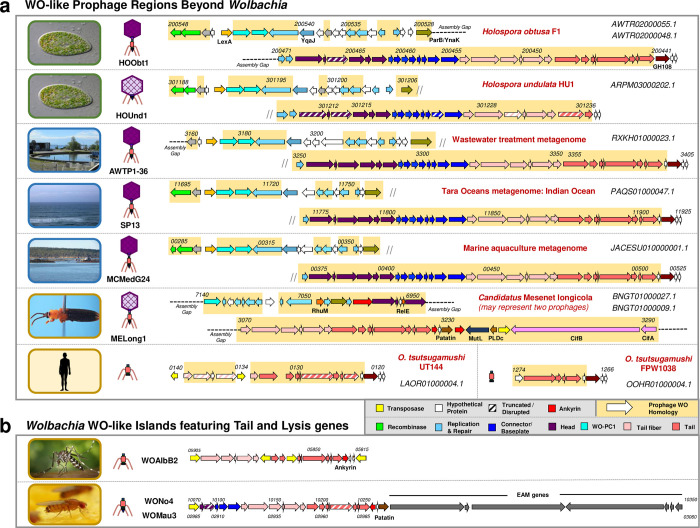
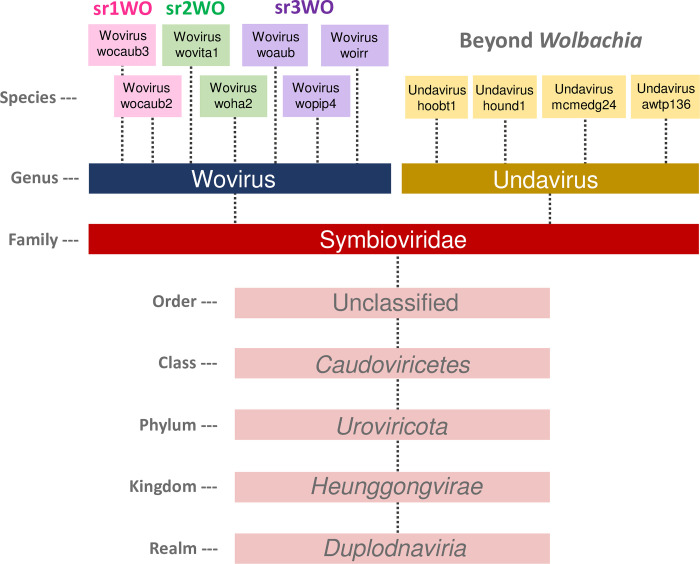
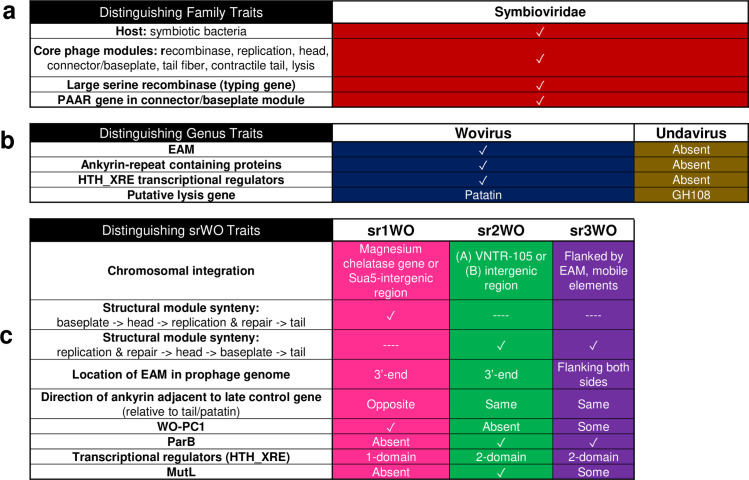
Wolbachia are the most common obligate, intracellular bacteria in animals. They exist worldwide in arthropod and nematode hosts in which they commonly act as reproductive parasites or mutualists, respectively. Bacteriophage WO, the largest of Wolbachia's mobile elements, includes reproductive parasitism genes, serves as a hotspot for genetic divergence and genomic rearrangement of the bacterial chromosome, and uniquely encodes a Eukaryotic Association Module with eukaryotic-like genes and an ensemble of putative host interaction genes. Despite WO's relevance to genome evolution, selfish genetics, and symbiotic applications, relatively little is known about its origin, host range, diversification, and taxonomic classification. Here we analyze the most comprehensive set of 150 Wolbachia and phage WO assemblies to provide a framework for discretely organizing and naming integrated phage WO genomes. We demonstrate that WO is principally in arthropod Wolbachia with relatives in diverse endosymbionts and metagenomes, organized into four variants related by gene synteny, often oriented opposite the putative origin of replication in the Wolbachia chromosome, and the large serine recombinase is an ideal typing tool to distinguish the four variants. We identify a novel, putative lytic cassette and WO's association with a conserved eleven gene island, termed Undecim Cluster, that is enriched with virulence-like genes. Finally, we evaluate WO-like Islands in the Wolbachia genome and discuss a new model in which Octomom, a notable WO-like Island, arose from a split with WO. Together, these findings establish the first comprehensive Linnaean taxonomic classification of endosymbiont phages, including non-Wolbachia phages from aquatic environments, that includes a new family and two new genera to capture the collective relatedness of these viruses.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
