

Pertussis toxin suppresses dendritic cell-mediated delivery of B. pertussis into lung-draining lymph nodes
- PMID: 35666769
- PMCID: PMC9216613
- DOI: 10.1371/journal.ppat.1010577
Pertussis toxin suppresses dendritic cell-mediated delivery of B. pertussis into lung-draining lymph nodes
Abstract
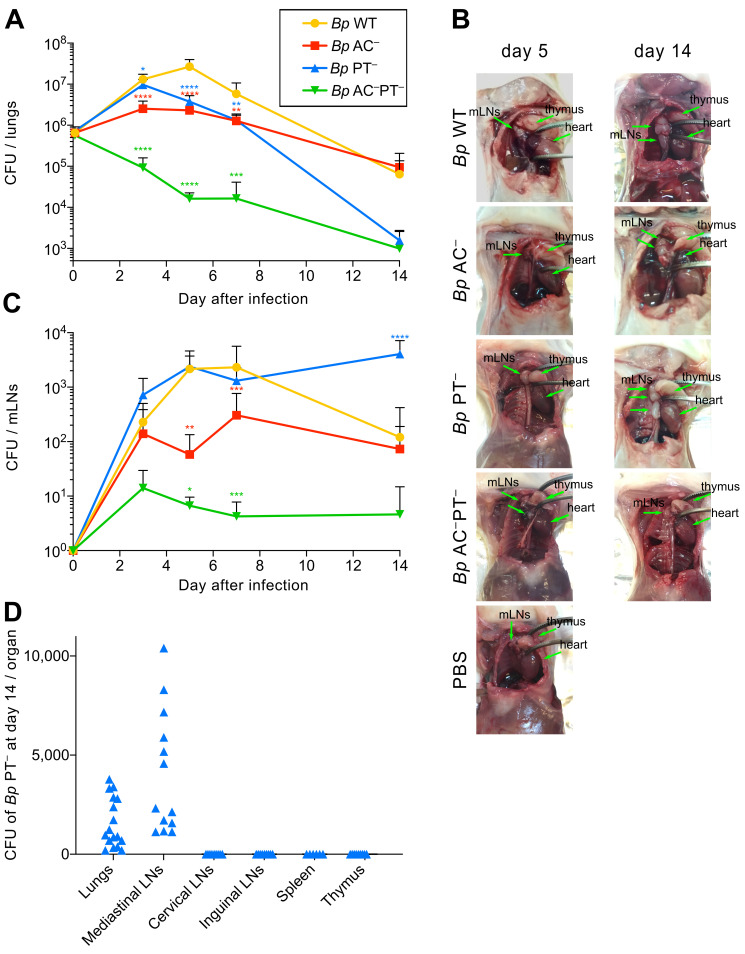
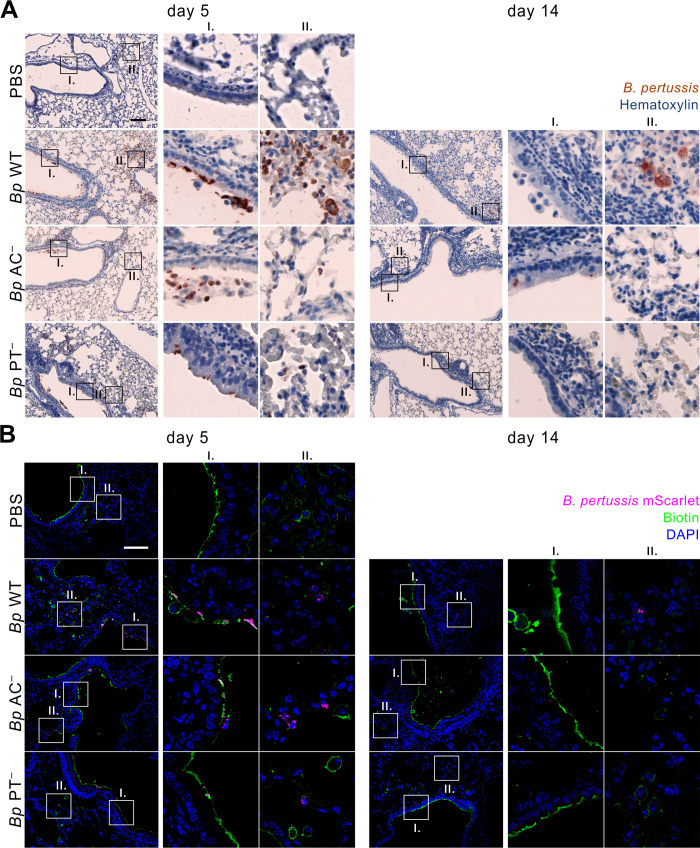
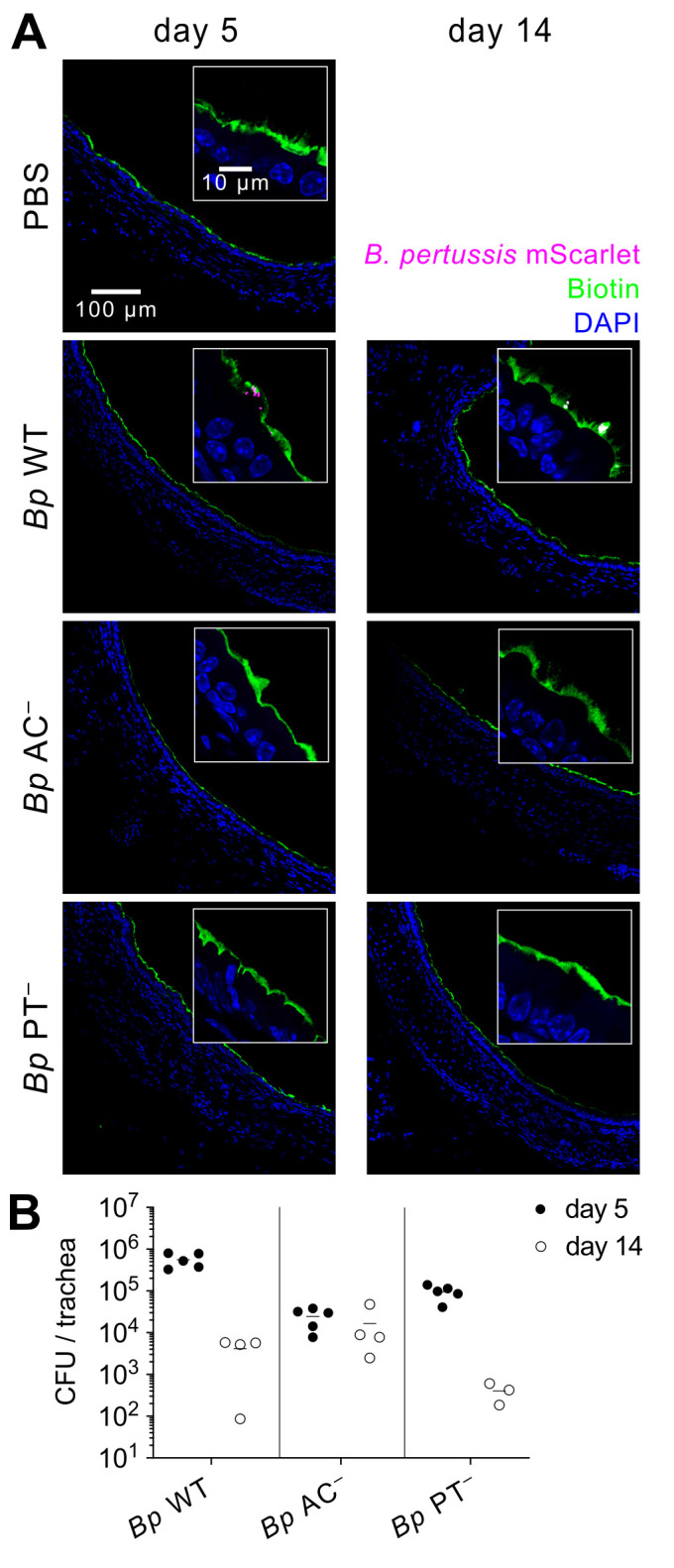
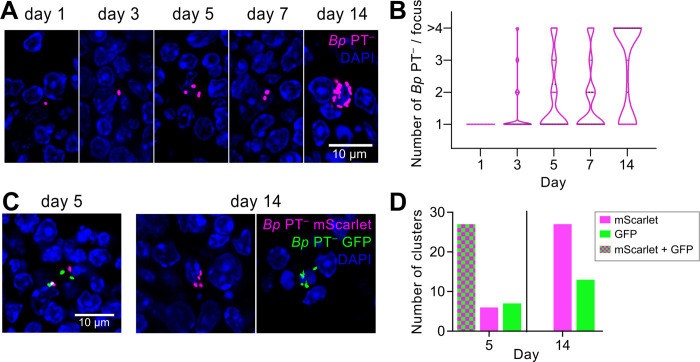
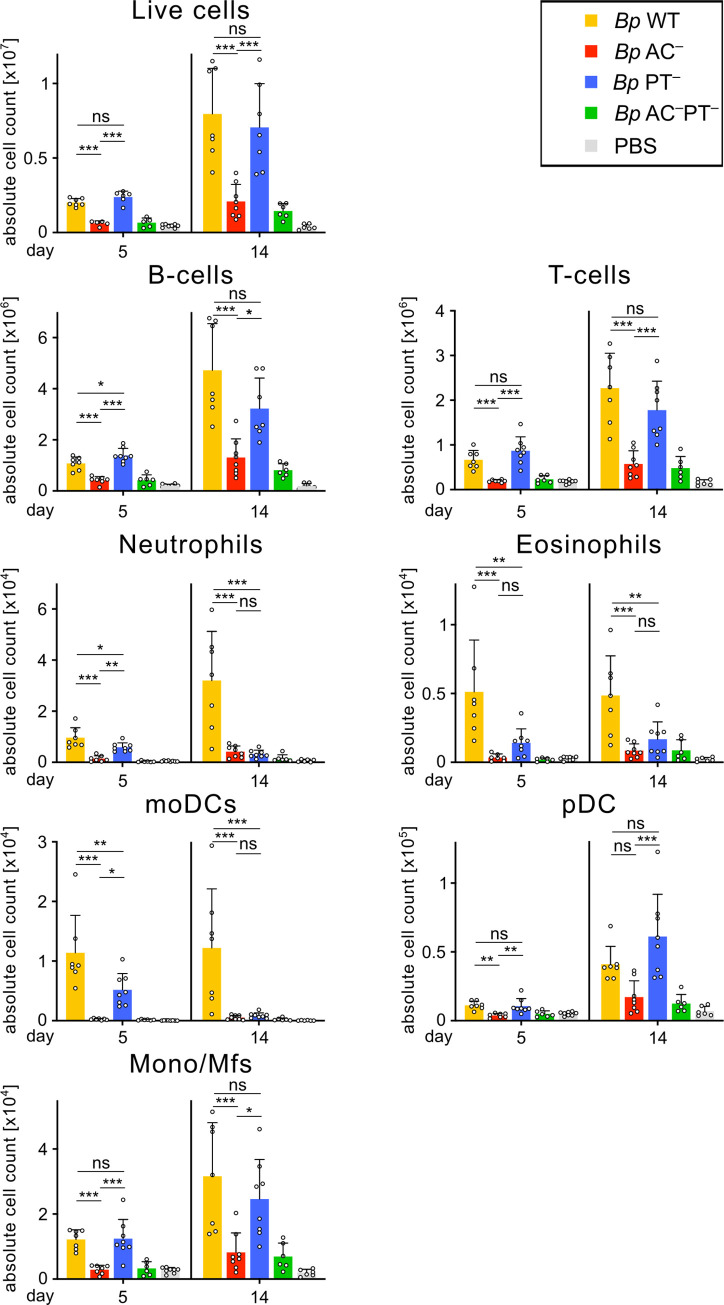
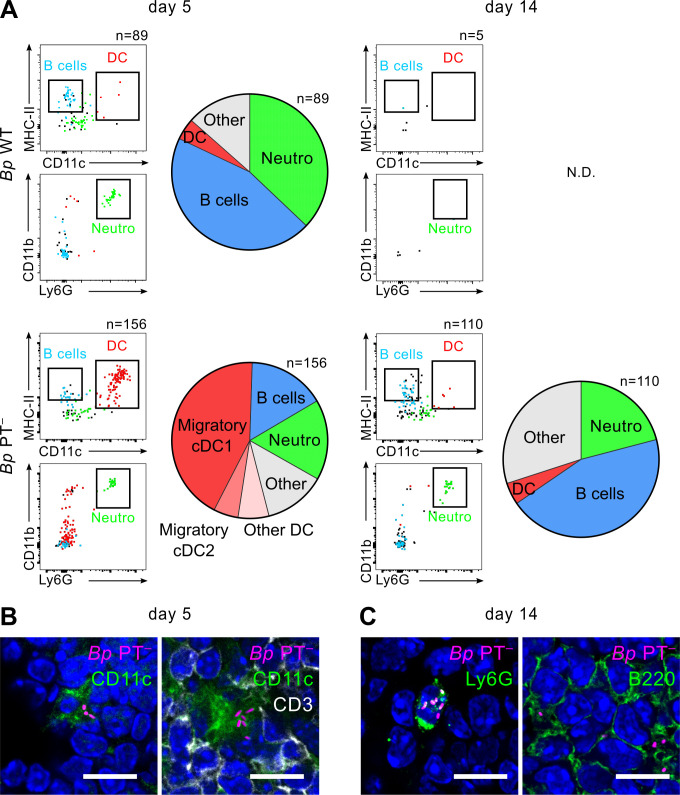
The adenylate cyclase (ACT) and the pertussis (PT) toxins of Bordetella pertussis exert potent immunomodulatory activities that synergize to suppress host defense in the course of whooping cough pathogenesis. We compared the mouse lung infection capacities of B. pertussis (Bp) mutants (Bp AC- or Bp PT-) producing enzymatically inactive toxoids and confirm that ACT action is required for maximal bacterial proliferation in the first days of infection, whereas PT action is crucial for persistence of B. pertussis in mouse lungs. Despite accelerated and near complete clearance from the lungs by day 14 of infection, the PT- bacteria accumulated within the lymphoid tissue of lung-draining mediastinal lymph nodes (mLNs). In contrast, the wild type or AC- bacteria colonized the lungs but did not enter into mLNs. Lung infection by the PT- mutant triggered an early arrival of migratory conventional dendritic cells with associated bacteria into mLNs, where the PT- bacteria entered the T cell-rich paracortex of mLNs by day 5 and proliferated in clusters within the B-cell zone (cortex) of mLNs by day 14, being eventually phagocytosed by infiltrating neutrophils. Finally, only infection by the PT- bacteria triggered an early production of anti-B. pertussis serum IgG antibodies already within 14 days of infection. These results reveal that action of the pertussis toxin blocks DC-mediated delivery of B. pertussis bacteria into mLNs and prevents bacterial colonization of mLNs, thus hampering early adaptive immune response to B. pertussis infection.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
